2022
261
19032021
16102021
 Olivera, J. M. juanmaolivera6@gmail.com
Olivera, J. M. juanmaolivera6@gmail.com
Ministerio
de Ganadería, Agricultura y Pesca, Uruguay
Riaño, M. E.
Universidad
de la República, Uruguay
Abstract: The invasion of ecosystems by exotic species has been identified as the second cause of biodiversity loss worldwide, and is one of the most difficult threats to reverse. In Uruguay, the introduction and spread of invasive alien species (IAS) has been identified as a serious environmental problem, becoming perhaps the greatest danger that native forests currently face. IAS often represents optical differences in the forest canopy and can therefore be detected remotely. The two most widespread and aggressive woody IAS in the country's forests are Ligustrum lucidum and Gleditsia triacanthos. The objective of this study was to spatially identify IAS within the native forest of Uruguay, mainly these two species, using remote sensing techniques. This work is based on multispectral data from medium-resolution satellite images (Landsat) and uses the normalized difference fraction index (NDFI) for classification. The NDFI is sensitive to canopy coverage and is calculated through a sub-pixel spectral mixture analysis (SMA), decomposing the reflectance information for each pixel into fractions. The results showed an area of 22,009 ha of native forest invaded by these IAS, with an overall accuracy of 87.6%, representing 2.63% of the total native forest area in the country. The results presented in this work will help to geographically analyze the invasion by IAS in the forest, linking it to possible drivers. Furthermore, this map can now be used as relevant information when designing IAS prevention, mitigation, restoration, and eventual eradication strategies in the country.
Keywords: forest degradation,Gleditsia triacanthos,biologic invasions,Landsat,Ligustrum lucidum.
Resumen: La invasión de ecosistemas por especies exóticas se identifica como la segunda causa de pérdida de biodiversidad a nivel mundial y se ha convertido en una de las amenazas más difíciles de revertir. En Uruguay, la introducción y la propagación de especies exóticas invasoras (EEI) son identificados como graves problemas ambientales, convirtiéndose quizás en el mayor peligro que enfrentan actualmente los bosques nativos. Las EEI a menudo representan diferencias ópticas en el dosel del bosque y, por lo tanto, pueden detectarse de forma remota. Las dos EEI leñosas más extendidas y agresivas en los bosques del país son Ligustrum lucidum y Gleditsia triacanthos. El objetivo fue identificar espacialmente las EEI del bosque nativo de Uruguay, principalmente estas dos especies, utilizando técnicas de teledetección. Este trabajo se basa en imágenes satelitales multiespectrales de resolución media (Landsat), utilizando el Índice de Diferencia de Fracción Normalizada (NDFI) para la clasificación. Este índice es sensible a la cobertura del dosel y se calcula mediante un análisis de mezcla espectral de subpíxeles (SMA), descomponiendo la información de reflectancia de cada píxel en fracciones. Los resultados mostraron un área de 22009 ha de bosque invadido por estas EEI, con una exactitud global de 87,6%, representando un 2,63% del área de bosque nativo. Este trabajo ayuda a analizar geográficamente la invasión de EEI en el bosque, relacionándola con posibles causales. Además, este mapa puede ser utilizado como información relevante a la hora de diseñar estrategias de prevención, mitigación, restauración y eventual erradicación de estas EEI.
Palabras clave: degradación de bosques, Gleditsia triacanthos, invasiones biológicas, Landsat, Ligustrum lucidum.
Resumo: A invasão de ecossistemas por espécies exóticas é apontada como a segunda causa de perda de biodiversidade no mundo e se tornou uma das ameaças mais difíceis de reverter. No Uruguai, a introdução e disseminação de espécies exóticas invasoras (EEI) é identificada como um sério problema ambiental, tornando-se talvez o maior perigo que as florestas nativas enfrentam atualmente. As IAS geralmente representam diferenças ópticas no dossel da floresta e, portanto, podem ser detectados remotamente. As duas EEI lenhosas mais difundidas e agressivas nas florestas do país são Ligustrum lucidum e Gleditsia triacanthos. O objetivo foi identificar espacialmente os EEI da floresta nativa do Uruguai, principalmente essas duas espécies, utilizando técnicas de sensoriamento remoto. Este trabalho é baseado em imagens de satélite multiespectrais de média resolução (Landsat), utilizando o Índice de Diferença de Fração Normalizado (NDFI) para classificação. Este índice é sensível à cobertura do dossel e é calculado usando análise de mistura espectral de subpixel (SMA), decompondo as informações de refletância de cada pixel em frações. Os resultados mostraram uma área de 22009 ha de floresta invadida por essas IAS, com uma precisão global de 87,6%, representando 2,63% da área de floresta nativa. Este trabalho ajuda a analisar geograficamente a invasão de EEI na floresta, relacionando-a com possíveis causas. Além disso, este mapa pode ser usado como informação relevante no desenho de estratégias de prevenção, mitigação, restauração e eventual erradicação dessas EEI.
Palavras-chave: degradação florestal, Gleditsia triacanthos, invasões biológicas, Landsat, Ligustrum lucidum.
Natural and environmental resources
Remote sensing of invasive alien woody species in Uruguayan native forests
Detección remota de especies leñosas exóticas invasoras en los bosques nativos de Uruguay
Detecção remota de espécies lenhosas exóticas invasoras nas florestas nativas do Uruguai
Olivera, J. M. juanmaolivera6@gmail.com
Riaño, M. E.
Received: 19 March 2021
Accepted: 16 October 2021
Published: 16 February 2022
Corresponding author: juanmaolivera6@gmail.com

The invasion of ecosystems by exotic species is the second cause of biodiversity loss worldwide, surpassed only by the destruction of habitats(1). These species cause environmental disturbances, including changes in the composition, structure, and processes of ecosystems, they can generate economic, social, and cultural problems, as well as problems in human and animal health(2)(3)(4). In Uruguay, the introduction and expansion of invasive alien species (IAS) has been identified as a serious environmental problem, particularly for biodiversity, becoming perhaps the greatest danger that native forests currently face(5).
In Uruguay, this process most likely has its origins in commercial and ornamental plantations near the native forest, which seeds are disseminated by different natural agents, making it a highly conducive environment for its growth and propagation(6). The conditions of forest soils, rich in organic matter and adequate levels of humidity, together with the protection against climatic agents and the action of livestock, provide a favorable environment for the development of IAS(6)(7).
Although some authors estimate that Uruguayan forests have increased in area in the beginning of the 21st century, an important degradation process has coincided with this increase(8)(9), particularly in the riparian forests of the southern and western coastal regions(10).
With the growing relevance of this issue, there is a need to advance research that allows for a clear diagnosis of the current situation of IAS, its territorial extension, its potential expansion, its impact on the integrity of forest ecosystems, and its ecological and socio-economic effects. To adequately manage forests and their services, via efforts in monitoring deforestation, special attention should be paid to the process of forest degradation caused by IAS in Uruguay.
Due to its continuity in time and its complete and consistent spatial coverage, satellite data provide valuable information, complementary to site-based measurements, for the detection of forest degradation processes(11). Some IAS invasions are subtle, occurring on a small scale or in the understory, and are therefore difficult to detect using satellite data. However, many invasive tree species can be perceived in the forest canopy, even dominating it, and can be remotely mapped(12). IAS often presents optical differences from native species and, therefore, can often be discriminated. The efficiency of detecting these species depends mainly on the sensor´s resolution (spatial, temporal, and spectral) and the differences (e.g. phenology, structure) between the IAS and nearby native vegetation.
The two most important invasive woody species in survey plots of the National Forest Inventory (NFI)(13) are Ligustrum lucidum (W.T. Aiton, Oleaceae) and Gleditsia triacanthos L. .Fabaceae), which will be the subject of this study.
Ligustrum lucidum or glossy privet is a woody species native to China and Japan, introduced in Uruguay in the mid-19th century(14). It is an evergreen species, unlike the native forest, with glossy privet with leaves all year round(15). It is associated with abundant ripe fruits (berries) throughout the winter and until the beginning of spring(16), a period in which the production of native fruits for birds decreases considerably. Its seeds have high germination power, fast growth, and can thrive under both sun and shade conditions(17).
These features, together with their abundant fruiting and wide tolerance to environmental conditions, allow it to establish itself as a dominant species in the forest, reducing or locally extinguishing the populations of some native species. Glossy privet invasion has been shown to limit native plant biodiversity and prevent the regeneration of native forests(18), possibly due to reduced light levels and soil fertility, with a high mortality of native seedlings(19). Thus, this species alters the structure, functionality, and composition of native ecosystems(20).
In Uruguay, glossy privet is one of the most invasive species with the greatest territorial significance(21), and is one of the 100 most damaging invasive species on a global scale(22). Its abundant presence has been borne out in the field, particularly in highly disturbed sites, such as those located on the sides of the bridges(23). However, there are no known records of when its expansion began, nor information regarding the drivers of it(21). Disturbances in forests can often increase the spread of this species or drive its invasion into new environments, resulting in new distribution patterns(24).
Gleditsia triacanthos or honey locust is a species with one of the greatest potentials to affect native Uruguayan ecosystems(7). It is a deciduous species native to the south-central region of the United States, which blooms in spring in the southern hemisphere, bearing fruit from early summer until late autumn(25).
In Uruguay, it is categorized as a IAS with a wide distribution and adverse impact(7). Some of the characteristics that favor its rapid expansion are its high resistance to environmental variations, such as floods or droughts, its high tolerance to contamination and soil salinity, its adaptation to any type of soil, its high growth rate, clonal and sexual reproduction, short juvenile period, abundant seed production, high germination capacity, and almost complete absence of pests and associated diseases affecting it(26)(27).
The main dispersal avenues of its seeds are watercourses and cattle, which forage their fruits and facilitate their germination(28). Animals, particularly cattle, contribute to consolidating the invasion process of this species by transporting the seeds and increasing their viability, favoring germination via the process of scarification(29).
Honey locust inhabits a wide variety of environments, but is commonly found in fertile, moist soils near streams. In Uruguay, the riparian forest seems to constitute its preferential habitat(30), although its spread includes prairie environments, marshes, and grasslands, lending it to an almost total replacement of native species(31).
Despite extensive experience in IAS control and advances in modelling their distribution patterns, there is a persistent lack of quantified and spatially explicit information on invaded forests in the Uruguay. Thus far, the majority of studies have used field data, and very few are based on remote sensing data. Some works in the region can be highlighted, such as the one carried out in Tucumán, Argentina, which was based on satellite images and in which differences in the phenology and productivity of forests invaded by glossy privet were observed. These presented higher NDVI values than nearby native forests, mainly because of the differences in the near-infrared band(32)(33). In a study carried out in the Yungas of north-eastern Argentina, higher values of NDVI (8.5%) were also observed in glossy private forests than in native forests(34). Hoyos and others(18) also used Landsat TM images to map areas of native forest invaded by glossy privet in an area of Córdoba, Argentina, using training points for classification based on the NDVI index(18).
The objective of the present study was to spatially identify IAS within the native forest of Uruguay, mainly glossy privet and honey locust, using remote sensing techniques. The goal was to detect glossy privet and honey locust due to their nature as widespread and aggressive IAS tree in Uruguayan forests(7), and the two most frequently observed IAS at the sampling points of the NFI(13).
To this end, we proposed classifying the country's forest into two classes –invaded and not invaded by the two prioritized IAS–, based on medium-resolution satellite images, evaluating the normalized difference fraction index(35) potential to reliably detect forest degradation by IAS. This methodology must be replicable over time to monitor its evolution over a given period.
The importance of this work lies in the usefulness of the generated spatial information for monitoring these species. Quantifying this invasion and obtaining a spatially explicit product at a national scale is an important tool to identify and prioritize areas for the control of these species, as well as to relate its evolution and distribution to possible drivers. A better understanding of these causes could be essential to identify the forests or sites at greater risk of being invaded by IAS. In this way, through appropriate forest management, the control and eventual eradication of these species could be more efficient, preventing the contamination of the most pristine forests and achieving a better conservation of forest biodiversity.
The Uruguayan woody flora is strongly influenced by neighboring provinces, especially from the sub-tropical Paranaense and Chaqueña floras(36). Based on the physiographic characteristics of the environments and their predominant species, the most widely used classification of native forest types in Uruguay in the literature(8)(10) is as follows: riparian forest, hillside forest, ravine forest, open forests, palm groves, psamophilic forests, and swamp forests.
This work was performed for the entirety extension of Uruguay. With a 173196 km. territory, this country is located between parallels 30° and 35° south latitude, and meridians 53° and 58° west longitude. The country’s climate is classified as a subtropical humid climate, temperate, rainy, and rainfall throughout the year, according to the Koppen climate classification. Uruguay is located in a transition biogeographic zone between different climatic areas called Pampean Province(37), characterized by a subtropical grassland matrix that is strongly modified by agroforestry and livestock activities. According to the national native forest cartography for 2016, it has an area of 835349 ha of native forest, almost 4.8% of the country's total area(38).
The national native forest cartography for 2016, produced by Sentinel-2 data classification, was used as a baseline in the construction of the IAS map, aiming to mask areas without native forest cover(38).
Data from the sampling plots of the NFI were used for the accuracy assessment. Of the total forest plots surveyed in the NFI (1490), 283 plots were registered with the presence of at least one alien species identified in the plot or its surroundings (18.99%). From the surveyed plots, 199 showed some individuals of Ligustrum sp. or Gleditsia triacanthos, corresponding to 13.35% of the total sampling plots.
In addition, to estimate the accuracy of the map after its final edition, georeferenced field records of IAS from the Biological Invasions Database for Uruguay(39) were used, along with field IAS records of REDD+ collaborating researchers. Another data source, the forest plantations map(40), was used to visually edit the map and eliminate errors in the algorithm with planted forests.
This work is based on surface reflectance data from the collections of the Landsat 5, 7, and 8 satellites, with multispectral sensors (TM, ETM +, and OLI), selecting images from Tier 1 (L1T), in which the USGS presents inter-calibrated data that already meets the geometric and radiometric quality requirements, so that no additional rectifications are required. These products contain a topographic correction for the displacement of the terrain due to relief. The only correction made was the masking of the cloudy pixels of the images, both with clouds and cloud shadows on the Google Earth Engine(41) platform.
The use of sub-pixel spectral mixture analysis (SMA), a technique based on the spectral modelling of images as a linear combination of pure spectra(42), has been a breakthrough for forest disturbance mapping. Monteiro and others(43) began to use these models for the detection of selective logging and fires in the Amazon forests of Brazil. Souza(44) proposed integrating the fractions of spectral endmembers of green vegetation (GV), soil, and non-photosynthetic vegetation (NPV) to map forest degradation(44).
Although several studies have been conducted on this subject, it is worth highlighting one by Souza and others(35), who proposed a new spectral index to improve the detection of forest canopy damaged by degradation, derived from selective logging and associated fires. This index is the normalized difference fraction index (NDFI), which is sensitive to canopy coverage and synthesizes reflectance information for each pixel, decomposed into the aforementioned fractions(35).
To calculate this index, an SMA is applied to the surface reflectace information from the multispectral image to decompose each pixel into sub-pixel purer material fractions or endmembers: GV, Soil, Shade, and NPV(42). The index is calculated as:

NDFI values range from ‒1 to 1, theoretically being higher in dense, undisturbed forests, with high canopy coverage, given by a combination of high GV fraction and low soil, shade, and NPV fractions. High NDFI values occur when the fractions of the spectral signature corresponding to soil and NPV remain close to zero, as a product of a completely covered canopy. The other two fractions fluctuate with seasonality, with the Shade fraction being higher in winter and lower in summer. On the other hand, the GV fraction generally presents high values in summer and low values in winter.
Based on this index, Bullock and others(45) used an SMA on Landsat time-series data to detect changes in forest cover, both deforestation and degradation in the state of Rondonia, Brazil(45). This method was applied using the Google Earth Engine(41) tool. Based on this research and the resulting algorithms, we proposed the detection of forest degradation using the NDFI for the classification of IAS.
As an example, the graphs below (Figure 1) show the evolution of NDFI over seven years (January 2010 to December 2017) in sites with no presence of IAS (A), extreme invasion of glossy privet (B), and extreme invasion of honey locust (C). This is based on data from NFI sampling plots and high-resolution complementary images. Some seasonal behavior of the index can be observed in A, most likely due to leaves falling (autumn-winter), and taking higher values in general during spring and summer. Forests in Uruguay can be classified as seasonal semi-deciduous, characterized by the senescence of 30-60% of the leaf biomass during autumn and winter(46), clearly differentiating itself from the glossy privet that maintains its foliage throughout the year.
Observing the evolution of NDFI in a forest with a ground certainty of invasion by glossy privet, the values were very close or equal to 1 throughout the year, except for some specific events. Similar behavior of the NDFI was observed in the data from adult commercial plantations of exotic species. For this reason, we decided to create the map using masking through a native forest layer to eliminate false positives and any confusion with commercial plantations. The same occurred to some extent in areas severely invaded by honey locust. However, in this case, this occurred with a notorious seasonality, typical of the species, where NDFI values close to 1 can be observed during the spring-summer season, when there is greater leaf coverage. For this reason, it was necessary to filter from the collection of available images, namely those acquired on a date such that this species had high leaf coverage and, therefore, high NDFI (December 1 to March 1).
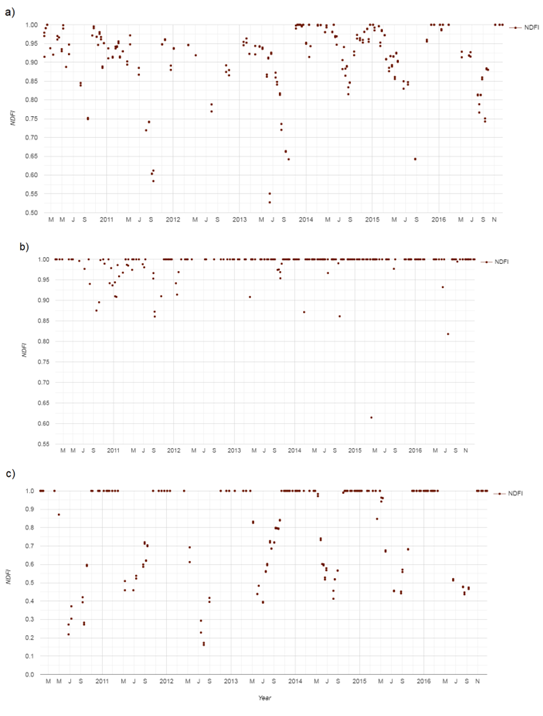
In summary, it can be observed that sites invaded by the two species studied behave very differently from the native forest with respect to this index. This may be due to the fact that these IAS form a denser and more homogeneous canopy than native forests, completely covering the ground and therefore reducing the soil fraction.
It is necessary to assume an operational definition of the degree of invasion that is supposed to be detected due to the limitations of the sensor (Landsat). Because of the spatial resolution of the images (30 m), the sensor is likely to detect highly invaded forest sites or IAS-dominated stands. In contrast, it has difficulty detecting sites with the presence of few IAS individuals or understory invasion. In this case, the resulting map would not correspond to a map of the absolute presence of IAS in Uruguayan forests.
For the generation of the IAS map, Google Earth Engine(41) was used to capture forest areas with high NDFI values throughout the country, adapting a script developed by Bullock and others(45), originally designed to detect disturbances in tropical forests. Certain considerations were considered:
- Landsat images were filtered by the acquisition date, using data from December 1 to March 1. The intention was to detect, in addition to glossy privet, areas dominated by honey locust or other deciduous exotic species, in the period of maximum leaf area, when NDFI was at its highest values.
- All Landsat images used were transformed into the mentioned endmember fractions using a script adopted from Bullock and others(45), which uses the “unmix” function in Google Earth Engine(41), unmixing each image pixel to return endmember proportions that sum to one.
- The calculation of the NDFI was performed on a five-year image stack (2013-2018) for the mentioned dates, to exclude the occurrence of extreme events, such as floods, where the NDFI may decrease more than usual. For this, the median of each pixel was calculated for each of the endmembers (“GV”, “Shade”, “NPV”, “Soil”) during the selected dates. With these values, the average of the NDFI was calculated for each year, and an average was found for the five-year period.
- Pixels were mapped with an average of the previous calculation greater than 0.99. This threshold was defined empirically by analyzing graphs of the index for georeferenced sites with field certainty of severe invasion.
Using QGIS(47), the generated layer was clipped by the limits of the 2016 native forest map. Thus, most commission errors generated by the commercial plantations were discarded. Nevertheless, some confusion may arise as a result of certain plantations mapped by mistake in the native forest cartography (false positives). Moreover, invaded forest areas may have been omitted because they are not part of the forest map (false negatives).
The final edition process consisted in creating a grid to divide the country into quadrants, examining them in an orderly fashion, editing the map visually by eliminating pixels confused with exotic commercial plantations or shade and shelter plantations using the forest plantation layer(40). Some less relevant alien species with similar characteristics may also be mapped with this methodology (as part of the commission error).
The results will be presented by Department and by ecoregion to analyze the geographical distribution of these IAS in the country, along with an invasion heat map.
All maps built from image classification through remote sensing contain errors. These errors are unavoidable and are a product of many factors, including clouds, class similarity, and climate variability(48). The most widely used methods for accuracy assessment are based on a confusion matrix(49). The confusion matrix is a table where the results of the map classification are displayed, along with the classification observed in a reference sample. The classes considered are the pixels belonging to the coverage of the IAS map and its complement (No-IAS). We assumed that the reference classification had the best assessment of the ground condition, and the global classification accuracy was defined as the proportion of correctly mapped cases. User’s accuracy was a proportion of cases mapped as IAS that was actually IAS “on the ground”. The producer’s accuracy consisted in the proportion of cases of IAS on the ground, which was also mapped as IAS(50).
Cohen’s kappa coefficient(49) is another statistic based on the confusion matrix, measuring the difference between the observed map-reality agreement and that which could be expected simply by chance. The kappa coefficient for the two classes is calculated as follows:
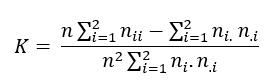
where is the total number of pixels in the map sample; is the number of observations in the main diagonal of the confusion matrix; are the marginal totals for the rows, and are the marginal totals of the table columns. The maximum and min-imum values of kappa depend on all the cells of the confusion matrix and not only on the values of the main diagonal; if all the elements outside the diago-nal are equal to zero, we would have complete agreement, and K = 1. The greater the agreement, the higher the value of the kappa coefficient.
Although the NFI sample was a systematic sample with national coverage, its sample design was not proposed for map accuracy assessment and may have limitations in making inferences. Furthermore, the selection mechanism for field samples corresponding to other data sources was unknown. To perform the accuracy analysis, we assumed that sample points were selected using simple random sampling. Thus, a confusion matrix was obtained directly from the sample.
Data not available: the data set that supports the results of this study is not publicly available.
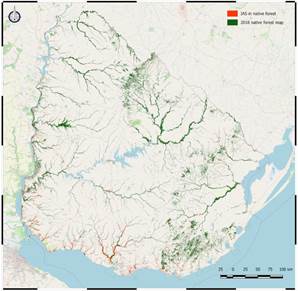
The following map (Figure 2) of the invasion of the studied IAS within the native forest was obtained for the period from 2013 to 2018 (highlighting its representation for better display), estimating a total area of 22008.9 ha of native forest with a high invasion of these IAS. This area represents 2.63% of the total 835349 ha of native forests in Uruguay(38).
Accuracy was assessed using a stratified reference sample, where the strata were map classes (IAS - No-IAS). From the NFI sample points, only plots where the severity of invasion of the species studied in this work was moderate, serious, or extreme, according to IUCN-CMP(13), were used, together with plots without any IAS to complete the reference sample. For the IAS class, 212 field observations were selected, and 488 for No-IAS strata. The sample of 212 points with IAS came from different sources (NFI, InBUy, and REDD+ Project collaborators). The 488 sample points belonging to the No-IAS strata were selected from the NFI (from a total of 1207 cases). Results are presented in Table 1.
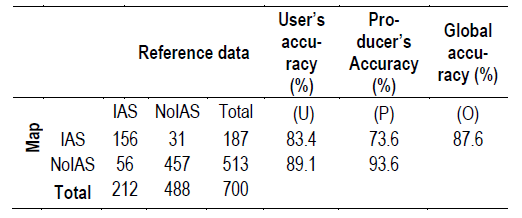
The global accuracy obtained was 87.6%, and the user’s and producer’s accuracy were 83.4% and 73.6%, respectively. The results show a high global accuracy (87.6%), as any value greater than 85% is considered a high value of accuracy(51). However, the kappa value obtained was 0.6955, and kappa values between 0.40 and 0.80 represent moderate agreement(52).
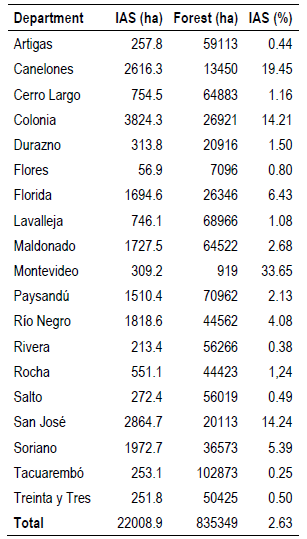
Table 2 shows the area occupied by the IAS and its percentage with respect to the total forest area, discriminated by department. Considering the percentages of invaded forests, it was observed that the most affected departments were Montevideo, Canelones, San José, and Colonia. On the other hand, the departments of Tacuarembó, Rivera, Artigas, and Salto have the lowest percentage of invaded forests.
This is consistent with the results obtained in the NFI, where the highest relative percentage of plots with IAS was recorded in the southern region of the country, where the departments of Colonia, Canelones, Montevideo, San José, and Soriano registered a relative percentage greater than 50%. In other words, in more than half of the plots surveyed for these departments IAS were registered(13).
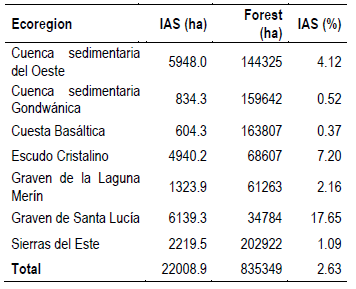
By analyzing the forest areas invaded by this IAS and its corresponding percentage for each ecoregion of the country(53), clear differences were observed among them. The highest percentage of invasion by ecoregion was observed in the Graven Santa Lucía region (17.65%), followed by the Escudo Cristalino region (7.20%), and in third place the Cuenca Sedimentaria del Oeste region (4.12%). In turn, the ecoregion with the least invasion corresponded to Cuesta Basáltica. This coincides with the information from the NFI, where a similar distribution was found, also highlighting the ecoregion of Graven Santa Lucía, where 68% of the sample plots had some IAS, followed by the Escudo Cristalino with 65%, and, third, Cuenca Sedimentaria del Oeste, with 36% of the surveyed plots (Table 3)(13).
Figure 3 shows a heat map of the presence for the two IAS studied, which can be considered a measure of the concentration of invaded forest sites. It is a density estimate (kernel regression) generated using QGIS(47). In this way, the heat map represents an index of the concentration of invaded forest polygons, weighted by the surface of these polygons.
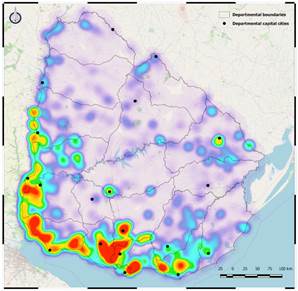
In the previous figure, the invasion of these two IAS is shown to have reached almost the entire territory to a greater or lesser extent. However, it also appears that the areas with the highest concentration were located on the west and south coast of the territory, mainly on the margins of the Río de la Plata and Uruguay rivers. On the contrary, we found that the northern region of the country was less affected by IAS than the rest of the country. Finally, some hot zones, mainly in the north, were found to be in the vicinity of important cities (e.g. Melo, Rivera, Salto, Paysandú, Durazno).
The approach proposed in this work represents the first useful and reliable tool for the mapping of degradation processes associated with IAS in Uruguay. This approach is characterized by the use of remote sensing techniques, and provides a basis for future works seeking to refine our results and detect other IASs.
According to the results obtained, a satisfactory level of accuracy was achieved, considering the complexity of detecting this type of process. The global accuracy obtained was 87.6%, which is comparable with the accuracy achieved in studies carried out in Argentina when mapping glossy privet invasion based on Landsat TM images using NDVI of 89%(18) and 84%(54).
This map helped to analyze the behavior of the most damaging IAS in the territory studied, linking them with possible drivers of degradation, and identifying and prioritizing areas for their control. Although there is a marked regionalization in the distribution of these IAS, they were found to have reached almost all the forests in the territory, establishing themselves as a real threat to the ecosystems found in the corresponding regions. Furthermore, our results indicate that the spread of IAS in the territory is even greater because this technique detects highly invaded areas, where these species are the dominant stands in the forest canopy.
The occurrence of hot zones near important cities, in terms of population, is reflected in the literature(18)(20)(54), where the link of the presence of IAS is pointed out, possibly due to the use of these species for ornamental purposes. For example, greater heat spots of glossy privet invasion were detected in the peri-urban areas of the largest cities in the northwest region of Argentina, where a relationship between the size of the cities and the area invaded by this species was observed(20).
A comprehensive analysis regarding what type, intensity, and extension of forest disturbances are those that favor the invasion of IAS in Uruguay is currently lacking(21). However, there is enough evidence in the literature to affirm that areas with a high degree of disturbance (e.g. land-use changes, selective logging, overgrazing, landscape fragmentation, urbanization, construction of bridges, and roads) generally have a higher degree of invasion, which could be mediating or enhancing the invasion process(24)(55)(56). Considering this, the information obtained could help to identify these disturbances as drivers of the invasion of IAS, linking its distribution with localized historical data of land-use changes and disturbances in the forest. It can be hypothesized that the high concentration of IAS in the south and west coasts of Uruguay may be linked to the fact that it is an area characterized by its high urbanization and intensive agricultural land use, which means a high degree of anthropization.
On the other hand, we found that the northern and northeastern regions had a lower presence of the prioritized IAS compared to the rest of the country. This may be due to various factors, including that forests in this area are further from the propagule sources marked as invasion heat zones (southwest); ravine forests that are present almost entirely in this area are generally less intervened forests and are difficult to access for livestock or for their exploitation. Therefore, they are generally in a better conservation state, reducing their vulnerability to invasions. Furthermore, these subtropical forests have greater biodiversity(10), a greater number of species per forest stratum, and a greater number of strata. Thus, the lower occurrence of invasion found in this region may be related to the hypothesis proposed by Elton(57), wherein the resistance of a biotic community to biological invasions increases in proportion to the richness of species present in the community(57).
Being able to spatially detect and quantify the invasion of IAS within the native forest is a key tool for defining prevention, monitoring, control, combat, and eradication strategies for invasive species. Analyzing the spatial distribution of the IAS, it was clear that there is a need to regionalize their management and control strategies, with different approaches and priorities depending on their location, severity of invasion, species involved, and valorization of the actually or potentially affected ecosystems.
Our findings allowed us to locate areas of potential IAS expansion due to their proximity to propagule sources, that is, sites with a high probability of being invaded in the future due to their proximity to areas already invaded and the pressure exerted by these propagules. This information can be used as an instrument for the implementation of strategies to prevent the entry of IAS into those sites. For example, considering the lower degree of invasion of the northern forests based on the results, strategies to prevent the entry of IAS could be implemented for this particular area. To this end, an initiative that instructs on the early detection of young individuals from prioritized IAS, stimulating their early extraction, could help prevent these invasions and keep the northern forests of the country in a better condition. A different approach should be proposed for the most affected areas of the south and west coast, where efforts should be focused on other objectives, in this case, prioritizing sites for the control and combat of these species. In any case, the proposed methodology can be used as a complement to the field data because it will probably be able to capture sites dominated by these IASs.
It is worth noting that, in most of the field data used for the evaluation, the degree of invasion was measured according to a relative criterion of presence of IAS, sometimes in the nearby area of the plot, and without having dasometric or phytosociological data of the IAS. Therefore, there may be cases in which a high degree of presence of IAS was registered in an NFI sample plot, but all these individuals were found in the understory, without being reflected in the canopy. In this case, the methodology may not be able to detect this invasion, forming part of the omission errors. Another situation arises with the date of NFI plots, which was carried out in three stages, 2009-2010, 2011, and 2014-2016(13). It may happen that at the time of NFI data collection, some plots did not show the presence of IAS and now they do. This situation also contributes to obtaining a higher omission error.
The use of medium-resolution satellite images (Landsat mission) to calculate the index NDFI has the potential to detect degradation in Uruguay´s native forests due to the invasion of alien species. In this work, it was possible to spatially identify areas dominated by IAS (at least for the two prioritized species, glossy privet and honey locust) with a satisfactory level of precision, considering the complexity of detecting this type of process. The classification obtained an overall accuracy of 87.6%, with an overall error of 12.4% and kappa coefficient of 0.695.
The quantitative and spatially explicit measure of these biological invasions acts as an effective indicator of forest degradation that is replicable in future studies to assess trends over time, considering the continuous monitoring of these invasions, as well as in reports at both national and international levels.
As such, this work helps to visualize and geographically analyze the invasion by IAS of the native forest, contributing to the knowledge of the distribution of these species. The results showed that an area of 22009 ha of native forest in Uruguay was invaded by IAS, which corresponds to 2.63% of the total native forest in the country.
In addition, the departments of the country most affected by these invasions were identified in this study, as well as the respective most affected ecoregions, thus improving the efficiency of possible invasion prevention, control, and mitigation actions of these invasive species. The highest concentration of sites invaded by IAS occurs in the southwestern region of the country, which is consistent with the available information collected in the NFI sampling plots(13); also coinciding with the presence of IAS in the Invasive Alien Species Database(39), as well as with much of the bibliography. The coastal departments of Montevideo, Canelones, San José, and Colonia are the ones with the highest percentage of IAS with respect to their forest area. On the other hand, the departments of Tacuarembó, Rivera, Artigas, and Salto were the least affected in terms of the percentage of forest invaded.
This map will help to identify priority sites of native forest for conservation, based on their ecological relevance and their invasion by IAS. In this way, proposals for local actions can be drawn up, focused on the conservation of certain sites, and on the mitigation or prevention of impacts derived from invasions by exotic species within the forest. The results of this study can also be used to design a IAS management plan at the national level and for possible ecological restoration actions at the site scale. Finally, further analysis of the information provided in this work could be used to identify possible drivers of species invasion.
Author contribution statement: JMO:
methodology design, image classification, mapping in geographic information
systems, analysis of results, interpretation of results, and article writing.
MER: contribution to data analysis,
accuracy assessment, and article writing.
Editor: The following editors approved this article. Gustavo Balmelli (https://orcid.org/0000-0002-5762-4207) Instituto Nacional de Investigación Agropecuaria (INIA),
Tacuarembó, Uruguay
http://agrocienciauruguay.uy/ojs/index.php/agrociencia/article/view/653/1089 (pdf)
This work was conducted within the framework of the REDD+ Uruguay project. We acknowledge the General Forestry Directorate (DGF) of the Ministry of Livestock, Agriculture and Fisheries (MGAP); the Biodiversity and Ecology of Conservation Group (BEC) of the University of the Republic, and the Invasive Species Database for Uruguay (InBUy) for making their field databases available.
juanmaolivera6@gmail.com


