1.
Introduction
The application of Integrated Pest Management programs is gaining more and more importance worldwide, as an approach that combines numerous sanitary techniques aiming to avoid economic damage caused by pests(1). The cornerstone of these programs is biological control(2), which is widely used in many countries and many situations, but has been developed especially in protected crops due to the high surface value of these productions(3). Control agents are periodically introduced to these crops, one or more times a year, so they can multiply in the crops, but without establishing permanently(4). In its implementation, several species of generalist predators are increasingly being released simultaneously against common pests(5)(6)(7). They are generally zoophytophagous predators, which is why they also include plant-provided food in their diet, but also other natural enemies(8)(9).
The species complementarity model suggests that herbivore mortality resulting from the combined action of different species of natural enemies is equal (additive) or higher (synergistic) than the sum of the mortality caused by each species of natural enemy(10). However, negative interactions between natural enemies, such as intraguild predation, facultative hyperparasitism, or interfering behavior, can affect herbivore suppression(11)(12). If negative interactions happen between control agents, rather than complementary effects, adding species can relieve phytophagous from the pressure of predation or parasitism(8). Negative interactions between natural enemies increase as diversity increases, making it necessary to decide between the release of a single superior species of natural enemies or multiple species against the herbivore pest(13). Intraguild predation is widespread in populations of natural enemies and occurs when one intraguild predator can negatively affect another, either by feeding on it or by competing for the shared resource (extraguild prey)(9). It can be unidirectional, with one species functioning as the top predator (i.e. intraguild) and the other as the intermediate predator (i.e. intraguild prey), or mutual, where the predators have the ability to consume each other(14)(15). Therefore, the combined release of generalist predators in these cases could have a negative result in the biological control of the target pest(16)(17).
In Uruguay, since 2013, numerous commercial producers carry out pest management of pepper cultivation under greenhouse based on the release of two predators, the mite Amblyseius swirskii (Athias) (Acarina: Phytoseiidae) used against whiteflies in tobacco, Bemisia tabaci (Gennadius) (Hemiptera: Aleyrodidae), and the flower bugOrius insidiosus (Say) (Hemíptera: Anthocoridae) for the control of western flower thrips, Frankliniella occidentalis (Pergande) (Thysanoptera: Thripidae). Such management is proposed as an alternative to the traditional one based solely on the application of chemical insecticides and it follows the guidelines of the usual practices in European countries(18) and in the United States(19).
Frankliniella occidentalis is a serious pest, vector of diseases (TSWV, Tomato spotted wilt virus) and the cause of significant damage due to feeding and oviposition(20). Its control with chemical insecticides is difficult, since they lay their eggs inside the plant tissues, and both adults and larvae feed in protected places, such as flower buds, which protect the pest from the action of insecticides(21). Orius insidiosus feeds on thrips larvae and adults(22); while A. swirskii mainly kills whitefly eggs and nymphs, but can also prey on first-stage larvae of thrips(23). Some studies indicate that Orius spp. could also feed on mites(18)(24). Therefore, although the generalist nature of Orius spp. can be an advantage when considering greenhouse pest control, it can also be a source of concern, due to the possibility of interference with other natural enemies(25). Such is the case of A. swirskii(26).
Due to this, there was a special interest in studying the compatibility of A. swirskii and O. insidiosus when used for the control of F. occidentalis, so as to know if their interaction increases or reduces the plague suppression. A second objective was to determine how prey availability may affect the tendency of O. insidiosus to kill A. swirskii. For this, the abundance of F. occidentalis was varied in experiments under controlled conditions.
2. Material and methods
2.1.
Experimental insects and plants
Colonies of F. occidentalis were established from individuals collected on pepper leaves and flowers (Capsicum annum L., Solanaceae) in the horticultural region of Salto (S 31°23′ - O 57°58′), Uruguay.
Thrips were multiplied in the Entomology laboratory at the Experimental Station of the Agronomy in Salto (EEFAS by its Spanish acronym) of the University of the Republic, following the protocol described by Steiner and Goodwin(27) and Espinosa and others(28). As plant substrate, bean seedlings were used (Phaseolus vulgaris L., Fabaceae), arranged in growth chambers in controlled conditions (25±1 ℃, 60±5% RH, 16:8 h L:D ). When the first leaves appeared, the petiole was cut off and placed in small glasses of water. The glasses were placed in glass containers (35 × 20 × 14 cm) with ventilated lids covered with fine mesh. Synchronized thrips cohorts were formed by placing adults on these leaves and transferring them to new leaves after 24 hours. These bean leaves containing the oviposition of the thrips were placed in boxes similar to those described above and kept for 6-7 days until the appearance of the second nymphal stage, or until 13-14 days when the adults appeared. These two states were used in the experiments. The individuals of A. swirskii and O. insidiosus were obtained from the Argentinian company Brometan. The A. swirskii mite was multiplied on pepper leaves, fed with eggs and nymphs of B. tabaci collected in commercial crops in Salto's horticultural region. The anthocorid O. insidiosus was reared using fresh bean pods as oviposition substrate and eggs of the flour moth, Ephestia kuehniella Zeller (Lepidoptera: Pyralidae) as prey, coming from the Production Unit of the Agronomy College. Each colony was placed in glass containers (35 × 20 × 14 cm) covered with low-density polyethylene (LDPE) stretch wrap with high transparency, in a climate-controlled chamber at 25 ± 1 ℃, 60 ± 5% RH 16:8 h (L: D).
Pepper plants (cvar Bilano type Lamuyo) and cowpea bean (Vigna unguiculata L. Walp.) grew individually in plastic pots of 20 cm in diameter and 10 cm deep, under controlled conditions (25 ℃ and 70% RH) with irrigation, in a polyethylene greenhouse of the Department of Plant Protection-Entomology at EEFAS, until they reached a suitable size for the experiments (20-30 cm in height).
2.2. Experimental unit and experiment
conditions
The
experiments with predators and thrips were carried out in an experimental unit
that included a pepper leaf disk (6 cm in diameter) placed on filter paper
moistened with distilled water. Said unit was placed in a 9.5 cm diameter x 1.5
cm deep Petri dish, vented through a hole in the lid covered with mesh (20 x 10
threads per cm2). During the 24-hour-long experiments, the plates
were kept at 25 ± 1 ℃,
60 ± 5% RH, 16: 8 h (L: D). O. insidiosus adult females were deprived of
food for 24 hours before their use.
2.2.1
Effect of O. insidiosus sex on A. swirskii and F. occidentalis
predation
To
evaluate possible differences in the predation of F. occidentalis and A.
swirskii by males and females of O. insidiosus, 10 adults of F.
occidentalis and 10 adults of A. swirskii were placed with a male or female of O. insidiosus in the
experimental units described above. Twelve repetitions were performed for each
sex of O. insidiosus. The number of dead prey
was counted 24 hours after the introduction of the anthocorids. To estimate
prey mortality due to manipulation or possible predation of F. occidentalis
by A. swirskii, 12 repetitions were carried out without the presence of O.
insidiosus.
2.2.2
Effect of different densities of F. occidentalis or A. swirskii
on predation by O. insidiosus
The predation capacity of O. insidiosus was evaluated on individuals of the second nymphal stage (6-7 days of life) and adults of F. occidentalis (13-14 days of life), and adults of A. swirskii, following the protocol described by Chow and others(26). Three densities were tested: 5, 10 and 20 preys (thrips or mites) per plate. Approximately one hour after the prey were placed, a single adult female of O. insidiosus was introduced. To avoid leaks, each plate was sealed with its lid and plastic wrap. At 24 hours, the number of dead preys inside the plates was counted.
Mortality due to handling was estimated from 10 repetitions for each type of prey (second stage or adult thrips) using 20 prey per plate without O. insidiosus. The handling was done with a fine tip brush.
2.2.3
Prey change by Orius insidiosus
Following the protocol established by Chow and others(26), the prey change of O. insidiosus females was evaluated by offering two combinations of prey: 5 F. occidentalis and 15 A. swirskii (1:3 ratio), 10 F. occidentalis and 10 A. swirskii (1:1 ratio), 15 F. occidentalis and 5 A swirskii (3:1 ratio). This experiment was repeated with second-stage nymphal and adult F. occidentalis individuals. In all cases, A. swirskii adults were used. Ten repetitions were completed for each relationship.
To do this, the required number of thrips was transferred to each plate, then the mites were incorporated and finally an O. insidiosus female. The dead preys were counted 24 hours after the introduction of the anthocorids.
The controls consisted of jointly depositing in the experimental units 20 F. occidentalis: 20 A. swirskii with no O. insidiosus. This experiment was conducted with second-stage nymphal and adult F. occidentalis individuals. In all cases, A. swirskii adults were used. Ten repetitions of each situation were performed.
2.3. Statistical analysis
All
counts and proportions were checked for normality and homogeneity of variance
(homoscedasticity). For the experiments with different prey densities, prey
change and choice with both sexes of O. insidiosus, a two-way ANOVA was
performed with “prey type” and “prey density” as factors to detect significant
differences in the total number of dead and recovered prey between treatments.
A one-way ANOVA was performed to compare the numbers of dead and recovered prey
in controls. Tukey's honestly significant difference (HSD) test was used to
determine significant differences between pairs of mean values after parametric
tests.
2.4. Data transparency
Available
Data: all the data set that supports the results of this study was published in
this same article.
3. Results
3.1. Effect of the O. insidiosus
sex on A. swirskii and F. occidentalis predation
The mean number of prey killed by O. insidiosus females was about 30% higher than the prey killed by males for the two prey types (p = 0.023). The numbers of thrips and mites killed by the O. insidiosus males were not different from each other (p = 0.756), while the females killed more thrips than mites (p = 0.038) (Figure 1).
Mortality of A. swiskii and F. occidentalis in the control plates was less than 10% of each type of prey and did not vary between the two types of prey (p = 0.842).
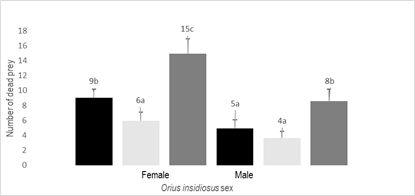 Figure 1
Mean number of dead prey (+SEM) of A.
swirskii and F. occidentalis adults per female and male of O.
insidiosus
Figure 1
Mean number of dead prey (+SEM) of A.
swirskii and F. occidentalis adults per female and male of O.
insidiosus
Black-F. occidentallis, Light
gray-A. swirskii, Dark gray-Total prey,
SEM: standard error of the mean
3.2. Effect of different densities of
F. occidentalis or A. swirskii on predation by O. insidiosus
Predation
was affected by the type of prey (p = 0.013) and the density of the
prey (p = 0.023). There was no interaction between prey type and density (p =
0.475). Predation of O. insidiosus increased with prey density (Figure
2). Prey mortality caused by the predator was higher than that registered in
the controls (p = 0.019). In these, the mortality of both thrips and mites was
less than 5% and did not vary with the prey type (p = 0.152).
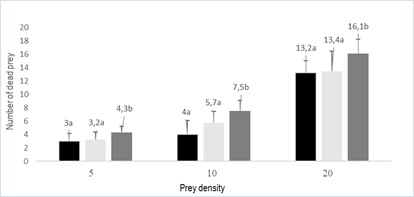 Figure 2
Mean number of dead prey of A. swirskii (adult) and F. occidentalis (2nd stage,
adult) (+SEM) by a female of O. insidiosus exposed to three prey
densities 5, 10 and 20 of each type
Figure 2
Mean number of dead prey of A. swirskii (adult) and F. occidentalis (2nd stage,
adult) (+SEM) by a female of O. insidiosus exposed to three prey
densities 5, 10 and 20 of each type
Black-A. swirskii adult, Light
gray- F. occidentalis adult, Dark gray-F. occidentalis nymph, SEM:
standard error of the mean
Predation
of F. occidentalis by O. insidiosus female increased with prey
density (p = 0.047), as well as the predation of A. swirskii adults (p =
0.032). No differences were found between F. occidentalis adults and A.
swirskii adults killed per Orius female (p = 0.613) at similar prey
densities. On the other hand, the predation of F. occidentalis nymphs
was greater than that of F. occidentalis and A. swirskii adults
(p = 0.035), which did not differentiate between themselves.
3.3. Change of prey by O. insidiosus
The
total predation of O. insidiosus was influenced by the type of prey, the
number of prey and the numerical relationship between the prey. The females of O.
insidiosus opted for the most abundant type of prey (p = 0.038), increasing
consumption when there was a greater number of prey available
(p = 0.045). When A. swirskii adults and F. occidentalis
adults were available in equal numbers, the females of O. insidiosus
showed no preference for one type of prey (p = 0.811) (Figure 3). However, when
the adults of A. swirskii and the second nymphal stage of F.
occidentalis were available in equal numbers, the females of O.
insidiosus killed twice as many thrips as mites (p = 0.013) (Figure 4).
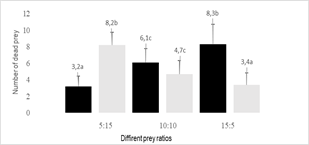 Figure 3
Average number of F.
occidentalis adults and A. swirskii adults killed (+SEM) by an O.
insidiosus female exposed to different numerical relationships between
preys
Figure 3
Average number of F.
occidentalis adults and A. swirskii adults killed (+SEM) by an O.
insidiosus female exposed to different numerical relationships between
preys
Black- F. occidentalis
adults, Gray- A. swirskii adults, SEM: standard error of the mean
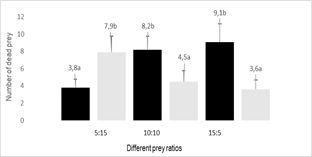 Figure 4
Average number of F. occidentalis nymphs and A.
swirskii adults killed (+SEM) by an O. insidiosus female exposed to
different numerical relationships between preys
Figure 4
Average number of F. occidentalis nymphs and A.
swirskii adults killed (+SEM) by an O. insidiosus female exposed to
different numerical relationships between preys
Black- F. occidentalis
nymphs, Gray- A. swirskii adults, SEM: standard error of the mean
The total number of dead prey was influenced by the numerical relationship between prey (p = 0.015). In all combinations, the most abundant prey was preferred (Figures 3 and 4).
Prey mortality was very low in the control treatments, and the number of dead prey of A. swirskii and adults or the second nymphal stage of F. occidentalis did not differ for each of the prey combinations (p = 0.620) and the average mortality ranged from 4 to 8%.
4. Discussion
The research studies that quantify the benefits of combining species of Orius and predatory mites for the control of F. occidentalis in greenhouse crops are few and their results vary since there are no differences between the release of a single agent compared to the release of both(26)(29)(30), to a greater reduction in the number of thrips if both agents are used simultaneously(31)(32).
In the present study, the intraguild predation model, which includes a higher predator (IG predator), an intermediate consumer (IG prey) and a shared resource(33), was confirmed in the interaction O. insidiosus - A. swirskii - F. occidentalis on a pepper leaf. Thus, Orius insidiosus killed the mite A. swirskii when it was available alone or in the presence of F. occidentalis. When the number of both preys was equal, O. insidiosus preferred the second nymphal stage of F. occidentalis over A. swirskii, and did not show differences between preys when considering adults of F. occidentalis and A. swirskii.
Our data show that the total and relative predation of F. occidentalis and/or A. swirskii by O. insidiosus depends on the type and number of prey available. When the number of individuals of the different types of prey was different, the preference in all cases was for the most abundant prey. Thus, when A. swirskii was the most abundant, that prey was preferred by O. insidiosus over F. occidentalis, which would indicate the possibility that O. insidiosus would replace the target prey, feeding on the other predator when thrips are scarce. Moreover, when A. swirskii is present in the crop, a significant increase in thrips would be necessary before O. insidiosus would prefer them. The statement by van Baalen and others(34) is reaffirmed, who state that changing prey can improve biological control by keeping predators on alternative prey when the target pests are rare, but it can also reduce control by distracting predators from attacking target pests, especially when they are not abundant.
These results demonstrate the need for caution when combining anthocorids and phytoseid mites for the control of F. occidentalis. Similar prey change patterns were found for O. insidiosus tested in combinations of A. degenerans and F. occidentalis(29), and of A. swirskii and F. occidentalis(26), both on greenhouse roses. This situation is also documented for other species of Orius against mites, such as O. majusculus(35), O. laevigatus(18)(24)(36) and O. tristicolor(25).
The results of this study were obtained by confining the two predators in a small plate. Caution is required when using small-scale laboratory experimental results to predict potential greenhouse interactions(37). Confrontations between O. insidiosus and A. swirskii are more likely in an experimental plate than in whole plants in a greenhouse. In addition, several factors may influence the intensity and outcome of intraguild predation, such as prey mobility, predation strategy, feeding specificity, relative prey size, and presence of extraguild prey(15)(37)(38)(39).
While much attention has been devoted to the influence of prey density and distribution on the feeding behavior of predators(40), these may also be influenced by the characteristics of the plants hosting their prey(41). Shipp and Whitfield(42) showed that the large number of trichomes in cucumber leaves, compared to pepper leaves, interfered with the search pattern of A. cucumeris (Oudemans). In addition, cutting leaves can affect the erection of the trichome and the chemical composition of the leaves, which in turn can influence the species observed(37).
It would be necessary to carry out complementary studies at the greenhouse level in order to clarify the final effect of using both predators together on the effectiveness of thrips control. According to Brodeur and others(43), intraguild predation by generalist predators is less important in greenhouses than in annual or perennial agroecosystems, since greenhouse crops are considered simple ecosystems with low biodiversity(44). The strength of the various interactions depends not only on the individual species involved but also on the suitability of the abiotic and biotic environment where the interactions take place. Interactions that appear very promising under controlled laboratory or greenhouse conditions have often been ineffective in the field due to unknown environmental constraints(45). Therefore, more research would be required for a better understanding of the interactions of change between anthochorids, predatory mites, and thrips in complex plant environments, and how these interactions would affect the suppression of thrips populations.
5. Conclusions
The
effect of intraguild predation between O. insidiosus and A. swirskii
requires caution if released simultaneously to control thrips F. occidentalis on
peppers in Uruguay.
Acknowledgments
We
are grateful to the authorities of the Experimental Station of the Agronomy
College (EEFAS by its Spanish acronym), and to the colleagues from the
Department of Vegetal Protection in EEFAS and the Entomology Unit in Montevideo
for their collaboration during the entire study period. This study was
supported by a grant from the National Research and Innovation Agency
(ANII/INNOVAGRO) that funded a project entitled “Contribution to the biological
control of pests of horticulture in the region of Salto.”
References
1. Viglianchino LE. Control integrado de Frankliniella occidentalis (Pergande) (Thysanoptera: Thripidae) con insecticidas y liberaciones de Orius insidiosus (Say) (Hemiptera: Anthocoridae) sobre pimiento en invernadero [grade’s thesis on Internet]. Santa Fé (AR): Universidad Nacional del Litoral, Facultad de Ciencias Agrarias; 2013 [cited 2021 Jul 07]. 76p. Available from: https://bit.ly/3yrCCzW.
2. van Lenteren JC. Controle de qualidade de agentes de controle biológico produzidos massalmente. In: Bueno VHP, editor. Controle biológico de pragas: produção massal e controle de qualidade. Lavras: UFLA; 2009. p. 311-37.
3. van Lenteren JC, Bueno VHP. Augmentative biological control of arthropods in Latin America. Biocontrol. 2003;48:123-39.
4. Jacas JA, Urbaneja A. Control Biológico de Plagas Agrícolas. España: Phtyoma Valencia; 2008. 496p.
5. van Lenteren JC, Bolckmans K, Kohl J, Ravensberg WJ, Urbaneja A. Biological control using invertebrates and microorganisms: plenty of new opportunities. BioControl. 2018;63:39-59.
6. Calvo FJ, Bolckmans K, Belda JE. Development of a biological control-based Integrated Pest Management method for Bemisia tabaci for protected sweet pepper crops. Entomol Exp Appl. 2009;133(1):9-18.
7. Shipp JL, Ramakers P. Biological control of thrips on vegetable crops. In: Heinz K, Van Driesche R, Parrella M, editors. Biocontrol in Protected Culture. Batavia: Ball Publishing; 2004. p. 265-76.
8. Rosenheim JA, Limburg DD, Colfer RG. Impact of generalist predators on a biological control agent, Chrysoperla carnea: direct observations. Ecol Appl. 1999;9:409-17.
9. Rosenheim JA, Kaya HK, Ehler LE, Marois JJ, Jaffee BN. Intraguild predation among biological-control agents: theory and evidence. Biol Control. 1995;5:303-35.
10. Snyder WE, Chang G, Prasad R. Biodiversity and successful conservation biological control: is there a relationship? In: Barbosa P, Castellano I, editors. Ecology of predator prey interactions. Oxford: Oxford University Press; 2005. p. 324-43.
11. Schmitz ОJ. Predator diversity and trophic interactions. Ecology. 2007;88:2415-26.
12. Rosenheim JA. Intraguild predation: new theoretical and empirical perspectives. Ecology. 2007;88:2679-80.
13. Bellows TS, Hassell MP. Theories and mechanisms of natural population regulation. In: Bellows TS, Fisher TW, editors. Handbook of Biological Control: Principles and Applications of Biological Control. Netherlands: Academic Publishers; 1999. p. 17-44.
14. Holt RD, Polis GA. A Theoretical Framework for Intraguild Predation. Am Nat. 1997;149(4):745-64.
15. Polis GN, Myers CN, Holt RD. The ecology and evolution of intraguild predation: potential competitors that eat each other. Annu Rev Ecol Evol Syst. 1989;20:297-330.
16. Rosenheim JA, Harmon JP. The influence of intraguild predation on the suppression of a shared prey population: an empirical reassessment. In: Brodeur J, Boivin G, editors.Trophic and Guild Interactions in Biological Control. New York: Springer; 2006. p. 1-20.
17. Janssen A, Willense E, van del Hammen T. Poor host plant quality causes omnivores to consume predator eggs. J Anim Ecol. 2003;72:478-83.
18. Urbaneja A, León FJ, Giménez A, Arán E, van der Blom J. Interacción de Neoseiulus (Amblyseius) cucumeris (Oudemans) (Aca.: Phytoseiidae) en la instalación de Orius laevigatus (Fieber) (Hem.: Anthocoridae) en invernaderos de pimiento. Bol San Veg Plagas. 2003;29:347-57.
19. Reitz SR, Yearby EL, Funderburk JE, Stavisky J, Momol MT, Olson SM. Integrated management tactics for Frankliniella thrips (Thysanoptera: Thripidae) in field- grown pepper. J Econ Entomol. 2003;96:1201-14.
20. Reitz SR. Biology an ecology of the western flower thrips (Thysanoptera: Thripidae): the making of a est. Fla Entomol. 2009;92(1):7-13.
21. Brødsgaard HF. Biological control of thrips on ornamental crops. In: Heinz KM, van Driessche RG, Parella MP, editors. Biocontrol in protected culture. Batavia: Ball Publishing; 2004. p. 253-64.
22. Simone M, Mendes E, Vanda H. Biologia de Orius insidiosus (Say) (Hemiptera: Anthocoridae) Alimentado con Caliothrips phaseoli (Hood) (Thysanoptera: Thripidae). Neotrop Entomol [Internet]. 2001 [cited 2021 Jul 07];30(3):423-8. Available from: https://bit.ly/3qTg1d7.
23. Wimmer D, Hoffmann D, Schausberger P. Prey suitability of western flower thrips, Frankliniella occidentalis, and onion thrips, Thrips tabaci, for the predatory mite Amblyseius swirskii. Biocontrol Sci Technol. 2008;18:541-50.
24. Lara L, van der Blom J, Urbaneja A. Instalación, distribución y eficacia de Orius laevigatus (Fiebre) y Orius albidipennis (Reuter) (Hemiptera: Anthocoridae) en invernaderos de pimiento en Almería. Bol San Veg Plagas. 2002;28:253-63.
25. Cloutier C, Johnson SG. Predation by Orius tristicolor (Hemiptera: Anthocoridae) on Phytoseiulus persimilis (Acarina: Phytoseiidae): testing for compatibility between biocontrol agents. Environ Entomol. 1993;22:477-82.
26. Chow A, Chau A, Heinz KM. Compatibility of Amblyseius (Typhlodromips) swirskii (Athias–Henriot) (Acari: Phytoseiidae) and Orius insidiosus (Hemiptera: Anthocoridae) for biological control of Frankliniella occidentalis (Thysanoptera: Thripidae) on roses. Biol Control. 2010;53:188-96.
27. Steiner M, Goodwin S. Methods for collecting and rearing thrips (Thysanoptera) and their natural enemies. Aust J Entomol. 1998;37:101-6.
28. Espinosa PJ, Fuentes JF, Contreras J, Bielza P, Lacasa A. Método de cría en masa de Frankliniella occidentalis (Pergande). Bol San Veg Plagas. 2002;28:385-90.
29. Chow A, Chau A, Heinz KM. Compatibility of Orius insidiosus (Hemiptera: Anthocoridae) with Amblyseius (Iphiseius) degenerans (Acari: Phytoseiidae) for control of Frankliniella occidentalis (Thysanopera: Thripidae) on greenhouse roses. Biol Control. 2008;44:259-70.
30. Skirvin DJ, Kravar-Garde L, Reynolds K, Jones J, Reynolds JJ, de Courcy Williams ME. The influence of pollen on combining predators to control Frankliniella occidentalis in ornamental chrysanthemum crops. Biocontrol Sci Technol. 2006;16:99-105.
31. Sörensson A, Nedstam B. Effect of Amblyseius cucumeris and Orius insidiosus on Frankliniella occidentalis in ornamentals. IOBC/WPRS Bull. 1993;16:129-32.
32. Ramakers PMJ. Coexistence of two thrips predators, the anthocorid Orius insidiosus and the phytoseiid Amblyseius cucumeris on sweet pepper. IOBC/WPRS Bull. 1993;16:133-6.
33. Holt RD, Huxel GR. Alternative prey and the dynamics of intraguild predation: theoretical perspectives. Ecology. 2007;88:2706-12.
34. van Baalen M, Krivan V, van Rijn PCJ, Sabelis MW. Alternative food, switching predators, and the persistence of predator-prey systems. Am Nat. 2001;157:512-24.
35. Brødsgaard HF, Enkegaard A. Intraguild predation between Orius majusculus (Reuter) (Hemiptera: Anthocoridae) and Iphiseius degenerans Berlese (Acarina: Phytoseiidae). IOBC-WPRS Bull. 2005;28(1):19-22.
36. Wittmann EJ, Leather SR. Compatibility of Orius laevigatus Fieber (Hemiptera: Anthocoridae) with Neoseiulus (Amblyseius) cucumeris Oudemans (Acari: Phytoseiidae) and Iphiseius (Amblyseius) degenerans Berlese (Acari: Phytoseiidae) in the biocontrol of Frankliniella occidentalis Pergande (Thysanoptera: Thripidae). Exp Appl Acarol. 1997;21:523-38.
37. Madadi H, Enkegaard A, Brødsgaard HF, Kharrazi-Pakdel A, Ashouri A, Mohaghegh-Neishabouri J. Orius albidipennis: intraguild predation of and prey preference for Neoseiulus cucumeris on different host plants. Entomol Fenn. 2008;19:32-40.
38. Madadi H, Enkegaard A, Brødsgaard HF, Kharrazi-Pakdel A, Ashouri A, Mohaghegh-Neishabouri J. Interactions between Orius albidipennis (Heteroptera: Anthocoridae) and Neoseiulus cucumeris (Acari: Phytoseiidae): Effects of host plants under microcosm condition. Biol Control. 2009;50:137-42.
39. Lucas E. Intraguild predation among aphidophagous predators. Eur J Entomol. 2005;102:351-64.
40. Luck RF. Principles of arthropod predation. In: Huffaker CB, Rabb RL, editors. New York: Wiley & Sons; 1985. p. 497-529.
41. Rapusas HR, Bottrell DG, Coll M. Intraspecific variations in chemical attraction of rice to insect predators. Biol Control. 1996;6:394-400.
42. Shipp JL, Whitfield GH. Functional response of the predatory mite. Amblyseius cucumeris (Acari: Phytoseiidae) on western flower thrips Frankliniella occidentalis (Thysanoptera: Thripidae). Environ Entomol. 1991;20(2):694-9.
43. Brodeur J, Cloutier C, Gillespie D. Higher-order predators in greenhouse systems. IOBC/WPRS Bull. 2002;25:33-6.
44. Enkegaard A, Brødsgaard HF. Biocontrol in protected crops: is lack of biodiversity a limiting factor? In: Eilenberg J, Hokkanen HMT, editors. Ecological and societal approach to biological control. Dordrecht: Springer; 2006. p. 91-122.
45. Heimpel GE, Mills NJ. Biological control: ecology and applications. New York: Chapman & Hall; 2017. 380p.
Author notes
meugenialorenzo@gmail.com
Additional information
Author contribution statement: MEL
collected the data and wrote the article, prepared and edited the figures. CB
contributed to the writing of some sections of the article and directed the
investigation. OL performed the statistical analysis of the data and was
co-director of the research. LB, GG and LM collaborated on the research
activities.
Editor: The
following editor approved this article.
Valentina Mujica (https://orcid.org/0000-0002-9820-2879)
Instituto
Nacional de Investigación Agropecuaria (INIA), Montevideo, Uruguay
Alternative link
http://agrocienciauruguay.uy/ojs/index.php/agrociencia/article/view/838/1004 (pdf)





