Introducción
El manglar es la asociación
ecológica de plantas arborescentes o arbustivas caracterizadas por adaptaciones
morfológicas y fisiológicas que les permite habitar zonas con la afluencia
periódica de aguas salobres. Por su naturaleza de selva anegada con agua salina
se le denomina halohelobioma. Este arreglo selvático marca la transición entre
el mar y la tierra, y es considerado en términos biológicos como la unidad
natural más productiva del planeta, permitiéndole por varios siglos ser un foco
de asentamientos y actividades humanas (Day et al. 1989; Prahl, 1989; Suman,
1994: Cantera y Arnaud, 1995). Esta situación, lo hace muy susceptible al
cambio, especialmente como consecuencia de actividades que el hombre ejerce
directamente sobre él para modificar, incrementar o disminuir el uso de sus
recursos (Botero y Mancera-Pineda, 1996).
Los manglares suramericanos se
distinguen de los manglares de los demás continentes tropicales por su escasa
diversidad de especies (Pannier y Pannier, 1976). Bajo los epítetos vernáculos
de mangle se agrupan varias especies disímiles, cuya única característica común
es crecer en suelos periódicamente anegados por aguas marinas.
Dentro del arbolado que puede
alcanzar 25-30 metros, se incluyen “mangle rojo” (Rhizophora mangle, R. harrisonii, R. brevistyla), “iguanero” o “salado” (Avicennia germinans), “piñuelo” (Pelliciera rhizophorae), “mangle blanco” o “mangle bobo” (Laguncularia racemosa) y “manglillo”,
“mangle zaragoza” o “mangle feliz”
(Conocarpus erectus). Este último no sobrepasa los 10 metros. A su vez
estos mangles se relacionan espacial y su- cesionalmente con el “nato” (Mora oleifera) (Sánchez-Páez, 1994).
Las comunidades de mangle manifiestan
patrones particulares de distribución dentro de su hábitat, aceptándose en
términos generales la siguiente zonación clásica a partir de la
línea de marea:
1. Mangle rojo, 2. Iguanero, 3. Mangle blanco, 4. Manglillo, 5. Piñuelo,
6. Nato (Prahl, 1989). Claro está que este patrón no siempre se mantiene, sino
que depende de una serie de factores y condiciones. Por lo que la distribución
está determinada por las distintas respuestas fisiológicas y morfológicas de
las plantas a los rangos mareales, al halogradiente, tipo de suelo y algunos
factores fisicoquímicos (Nickerson y Thibodeau, 1985; Prahl, 1989). La
alteración de las condiciones para su desarrollo por la presencia de uno o
varios factores de origen natural o antropogénico, puede obligar al sistema de manglares
a un mayor gasto energético para mantener la homeóstasis. Los agentes que
determinan esta condición son llamados tensores (Lugo et al. 1980; Ricaurte et
al. 1992).
Ancestralmente, las comunidades afrocolombianas han mantenido una relación estrecha con los recursos de los ecosistemas de manglar a lo largo de la franja del litoral pacífico, (Bravo-Pazmiño 1998), abasteciéndose de madera para diversas construcciones, utensilios domésticos y de pesca, combustible, obtención de sustancias medicina- les y también de su fauna asociada como fuente primaria de obtención de proteína animal (Prahl et al. 1990).
A partir de los años 50 la demanda en las ciudades del interior, estimuló la explotación industrial del manglar. El corte masivo de madera para construcción, obtención de pulpa de papel, fabricación de carbón vegetal y la extracción de taninos, fueron parte de una inmensa campaña extractivista sobre este ecosistema (Prahl et al. 1990). Las consecuencias en las características estructurales de la vegetación han puesto en pe- ligro al ecosistema de manglar y a su biocenosis (Sánchez-Páez et al. 1997).
Dentro de los estudios que relacionan los cambios en el manglar y la acción antrópica, se encuentra el de un manglar en Puerto Rico, en el cual Holdridge (1940) plantea que los desmontes dan origen a cambios en la composición, ya que conducen a cierta degradación de la masa arbórea y retrasan la sucesión en las primeras fases. En ese estudio el porcentaje de cada especie antes de la intervención humana era la siguiente: L. racemosa 30%, A. germinans 30%, R. mangle 30 %, C. erectus 10%; después de los desmontes respectivamente era: 50%, 25%, 20% y 5%. En Cuba áreas taladas hace 10 años ocupadas originalmente por A. germinans, hoy muestran gran regeneración de L. racemosa (Báez-Valdez y González-Rondón, 1980).
West (1977) citado por Bejarano et. al. (1992) establece para la costa pacífico que una “alta tasa de tala” disminuía la densidad total de las selvas y estaba asociada a una aparición de plantas invasoras de origen terrestre como Acrostrichum aureum. Ricaurte et al. (1992) y Blanco-Libreros y Cantera (1995) establecieron a su vez que la tala determina una disminución del DAP (diámetro a la altura del pecho) promedio de las selvas, por la aparición de nuevas plántulas y la pérdida de árboles de mayor porte; además la tala tiene un efecto sobre la dominancia de las especies en los manglares, puesto que cambia la composición de especies. En la Bahía de Buenaventura las selvas perturbadas por la tala presentan mayor dominancia de A. germinas y P. rhizophorae ya que se ejerce una presión selectiva sobre los árboles de Rhizophora spp., o de M. oleifera (Blanco-libreros y Cantera, 1995).
La dinámica ecológica del manglar plantea que este va colonizando cuerpos de agua gracias a las raíces zancudas de R. mangle (Mizrachi et al. 1980), la cual sirve de especie “terrígena”, esto es, fijadora de sedimentos que no solo consolidan amplios segmentos del litoral sino que con su acción el litoral progresivamente le va ganando terreno al mar (Hernandez-Camacho et al. 1980; Horna-Zapata, 1980). Las formaciones de A. germinans y L. racemosa tienden a su vez a colonizar el área ocupada por las formaciones más viejas de mangle rojo ya consolidadas. Esto muestra como la sucesión depende de la modificación de las condiciones ambientales por cada especie sucesiva. La modalidad de zonificación según Odum (1969) y Lanly y Lindquist (1985) refleja en gran medida la sucesión que tiene según va afirmándose el suelo, dándole al ecosistema manglar su naturaleza dinámica.
Sin embargo no siempre la zonación recapitula la sucesión. Esto depende de características propias de cada uno de los ecosistemas de manglar. La sucesión puede ser cíclica y guiada por una serie de estadios cíclicos y los cambios en cada estadio están controlados por uno o varios factores externos. Por lo tanto la sucesión en los manglares exhibe patrones que son típicos de los ecosistemas estresados, por ejemplo la autosucesión. La diversidad específica en estos casos puede crecer si hay una disminución en la intensidad del tensor, en cambio cuando el tensor antrópico se impone sobre los tensores naturales la complejidad estructural del manglar se reduce. A pesar de esto último, el manglar continúa reproduciéndose y manteniéndose en los sistemas costeros de baja energía y no es reemplazado por otros sistemas (Lugo, 1980).
Si bien los efectos de los tensores antrópicos sobre el manglar como la tala, se han estudiado (Holdridge, 1940; Báez-Valdez y González-Rondón, 1980; Bejarano, et al. 1992) dentro de estos trabajos no se ha abordado en su conjunto, la relación existente entre este ecosistema y las comunidades que lo habitan, poniéndose de manifiesto en el tiempo histórico, los cambios en su dinámica (zonación, estructura y geomorfología) y las causas naturales y antrópicas de estos.
Muchos trabajos han estudiado la respuesta del ecosistema de manglar ante alteraciones físicas como construcción de carreteras, diques y terraplenes (Hernández-Camacho et al. 1980; SánchezPáez, 1994; Serrano et al. 1995) o químicas como derrames de petróleo (Bohorques y Prada, 1984). Sin embargo, no se ha profundizado el impacto que pudiera causar la prolongada explotación de su madera sobre la estructura y zonación de las asociaciones vegetales (De Las Salas y Hildebrand 1980; Ricaurte et al. 1992) y sobre la geomorfología de los sistemas estuarinos.
Tratándose de un ecosistema de gran valor ecológico y comercial (Prahl, 1989), resulta muy importante establecer cuáles son los impactos que las actividades antrópicas ocasionan en su estructura y dinámica ecológica, por lo que esta investigación se planteó determinar la influencia de los impactos antrópicos en la estructura del manglar en la ensenada de Virudó, Bajo Baudó, Chocó, Colombia. Como subcomponentes se enfocaron varios ejes temáticos así: reconstruir la historia de uso y explotación que el recurso madera del ecosistema manglar ha tenido en la ensenada de Virudó; Determinar los volúmenes aproximados de extracción (tradicional e industrial), con los lugares y épocas de explotación, que las diferentes especies de mangle de la ensenada de Virudó han tenido y reconstruir la cronología de los cambios acontecidos en la estructura y zonificación del ecosistema manglar y en la dinámica costera de la ensenada (procesos de erosión y acrecimiento de playas y firmes, fluctuaciones en la línea costera), ubicando en la base cartográfica de la ensenada los lugares de estos cambios y el tiempo en el cual ocurrieron.
El marco hipotético plantearía que la dinámica (estructura, zonación y geomorfología) del ecosistema de manglar de la ensenada de Virudó se ha visto modificada en el tiempo por la acción que el hombre ha ejercido sobre él. Así mismo se plantean pregunta como ¿Existe una relación entre los cambios en la dinámica del ecosistema de manglar y los cambios en las formas de uso del ecosistema? ¿Hay diferencias en el impacto sobre el ecosistema entre extracción comercial y extracción artesanal? ¿Hay relación entre los cambios en la dinámica del manglar y la dinámica costera?
Metodología.
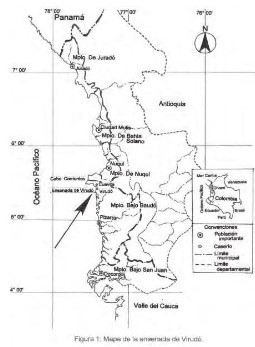
Área de Estudio. Corresponde a la ensenada de Virudó-Cuevita ubicada en la parte
norte del mu- nicipio del Bajo Baudó (Chocó), costa pacífica colombiana
(05.40248° N; 077.40664° W) (figura 1). Limita al norte con Cabo Corrientes, al
oriente con la serranía del Baudó, al occidente con el océano Pacífico y al sur
con la desembocadura del río Virudó. En esta área están ubicados los consejos
comunitarios de Cuevita y Virudó, quienes ejercen la mayoría de sus actividades
tradicionales y de subsistencia en el ecosistema de manglar que se forma en
esta ensenada por la afluencia de los ríos Virudó, Virica, Jella, Ocaba y
Pavasa. Estas comunidades pertenecen a la génesis afrocolombiana y habitan
estos territorios desde hace más de un siglo. La localidad de cuevita se
localiza en el extremo norte de la comba que forma la prolongación rocosa de
Cabo corrientes. Está conformada por 44 viviendas y una población de 188
habitantes que se distribuyen en 34 grupos familiares. Virudó es el
corregimiento más grande del área con 743 habitantes y se ubica en el extremo
norte de la isla de barrera de Virudó. Estas comunidades se dedican a la pesca
artesanal, a la agricultura y a la explotación maderera para la venta en los
aserríos locales que están en manos de personas foráneas (Bravo-Pazmiño, 1998).
 Figura
1.
Ubicación área de estudio
Figura
1.
Ubicación área de estudio
Zona de vida. Según el modelo de Holdridge se trata de Bosque pluvial Tropical; con una biotemperatura de 24°c, un promedio de precipitación que excede los 8000 mm al año y la humedad oscila entre 75 en época seca y 99% durante época lluviosa (Sánchez-Páez et al. 1997). Es de resaltar que esta ensenada posee el primer manglar de la llanura costera aluvial que empieza a formarse desde cabo Corrientes hacia el sur. Este manglar se puede considerar como el prototipo para esta zona media (hasta la desembocadura del río San Juan) y se caracteriza por un sistema de barras arenosas de protección y una red interna de esteros y canales mareales, dominados en gran parte por Rhizophora spp que lo convierten en un estuario complejo. De acuerdo a la zonificación preliminar hecha por el Ministerio del Medio Ambiente de los manglares del Pacífico se le considera como perteneciente a la Zona de Uso Múltiple de Virudó (Prahl et al. 1990; Sánchez-Páez et al. 1997).
La metodología del presente proyecto partió de presentar y discutir el proyecto ante la Asamblea General de los Consejos Comunitarios de Virudó y Cuevita. Es importante anotar que el acercamiento y el trabajo conjunto de la Fundación Iniciativa Baudó y estos Consejos Comunitarios durante los 5 años anteriores a la iniciación del presente proyecto permitieron tejer con los habitantes lazos afectivos y de confianza que redundaron en su interés por participar en el proyecto, aún sin que hubiera una compensación salarial, y en garantía de la veracidad de los testimonios. La reconstrucción histórica del uso, explotación y cambios del ecosistema de manglar comenzó con la elaboración de la cartografía social del ecosistema de manglar según Santamaría y Andrade (1997). Se realizó un taller en el consejo comunitario de Virudó con seis hombres mayores y nativos de los consejos comunitarios de Virudó y Cuevita. Se invitó a los mayores a que sobre la cartografía IGAC (Planchas 183-III-B y 183.III-D de 1988) ubicaran o verificaran los ríos, quebradas, esteros y asentamientos, al igual que las zonas ocupadas por distintas asociaciones vegetales del ecosistema de manglar y natal. Esta primera zonación del manglar se contrastó con el testimonio de otros miembros de ambas comunidades a quienes se les preguntó detalles sobre las zonas del manglar en las que han trabajado o que frecuentan por estar en la ruta de paso hacia su finca. Igualmente se hicieron recorridos, a canoa y en lancha a motor, por toda la ensenada, los ríos y esteros para verificación y corrección en casa de que fuera necesario. En estos recorridos se tomaron las coordenadas de los puntos donde se daba la transición de manglar a natal con un GPS (GARMIN GPS 38 FCC ID IPH-13001).
Sabiendo que la memoria es un elemento clave en el proceso de
reconstrucción de la experiencia humana, transportada y traducida a la
actualidad por los relatos de las fuentes de información o informantes,
(Aceves-Lozano, 1999) y que las Comunidades Negras del litoral Pacífico como
cultura privilegian la tradición oral; se siguió el método de investigación de
las historias orales temáticas según Acevez-Lozano (1999). Mediante entrevistas
abiertas a 25 personas mayores (anexo B), en su mayoría líderes de la
comunidad; se hizo la reconstrucción histórica de uso, explotación y cambios
(estructurales y geomorfológicos) del ecosistema de manglar. En las entrevistas
se indagó sobre la clasificación etnobotánica de las especies maderables del
manglar, usos, volúmenes requeridos y extraídos, zonas de explotación, épocas
de explotación, cambios en la estructura y zonación del manglar y en la
geomorfología del sistema estuarino de la ensenada (procesos de erosión y
acrecimiento y modificaciones en la línea costera). Por medio del procedimiento
directo se privilegió el testimonio de antiguos aserradores y los mayores que
tradicionalmente han usado el manglar para sus actividades. A pesar de
privilegiar la fuente oral se procedió a triangular o extrapolar las fuentes
constitutivas (Denzin, 1990, citado por AcevezLozano, 1999), es decir
convalidar la información a partir de fuentes documentales.
Este enfoque metodológico privilegió la participación comunitaria en la construcción del conocimiento o proceso de investigación. Esto parte del supuesto de que la comunidad, que ha vivido dentro del ecosistema de manglar y hace uso de él para su sostenimiento, conoce más que cualquier otro sujeto el entorno en que vive y los cambios que ha sufrido en el tiempo. Esta parte de la investigación correspondería a lo que Schutter (1980) plantea: “Producción de conocimientos sobre la relación dialéctica entre los sujetos y la objetividad, es decir, entre las estructuras objetivas y la manera en la que se perciben a sí mismos y la relación con estas estructuras”.
Para el estudio de la zonación y la estructura del ecosistema de manglar se realizaron 6 transectos de 200 metros de largo, perpendiculares a la orilla del estero en dirección al interior del bosque. Dos en el estero Bocón (transecto 1 N 05.39743° W 077.39761° y transecto 2 N 05.39294° W 077.40254), dos en el estero Peñitas (transecto 3 N 05.25102° W 077.22799 y transecto 4 N 05.42382° W 077.38532°) y dos en el estero El Encanto (transecto 5 N 05.42382° y transecto 6 N05.46512° W 077.42095° 077.42095°)(figura 2)
Criterios de ubicación de los transectos:
• Mayor área de estudio posible.
• Diferentes condiciones ambientales (influencia cercana al mar, influencia de los ríos).
• Mayor información recogida en la cartografía social e historias de vida.
En cada transecto se delimitaron desde la
orilla 10 parcelas de 100 m2 (0.01 ha) separadas entre sí por 10 m. En cada
parcela se cuantificaron los atributos de la vegetación de acuerdo con los
métodos de Schaeffer-Novelli y Cintrón (1986). Se consignó la siguiente
información: el diámetro a la altura del pecho (DAP) y la altura solamente a
los árboles vivos con diámetros superiores a 2.5 cm. En cada parcela se tomaron
5 muestras de sustrato de 25 cm x 25 cm x 25 cm de profundidad para hacer
análisis granulométrico de acuerdo con Horna-Zapata (1980). Adicionalmente, se
midió la salinidad intersticial en 5 puntos de cada parcela mediante un
salinómetro-refractómetro y se tomaron datos cualitativos sobre la vegetación y
la fauna asociada (presencia de algas, moluscos y epífitas).
Trabajo de Laboratorio. La cartografía social obtenida se digitalizó en AUTOCAD 14. Después de hacer el procesamiento de la información georreferenciada de GPS y otras fuentes geográficas se consignó esta información en la cartografía digitalizada. A partir de las fotografías aéreas obtenidas en el centro de documentación de INGEOMINAS-CALI (Plancha 183, sobre No. 23751, vuelo c-1224 de 1968, fotos 188-198, escala 1:40000; sobre No. 760, vuelo M-1087 de 1961, foto 1557, escala 1:65000) se hizo la fotointerpretación de la cobertura vegetal del ecosistema de manglar con un estereoscopio de espejos WILD HEERBRUGG. Los criterios utilizados para la fotointerpretación fueron los siguientes: textura del dosel, forma de las copas y altura del arbolado. En la fotointerpretación también se ubicó zonas intervenidas, asentamientos y aserríos. Los mapas obtenidos se transformaron a escala 1:25000 mediante un STEREO ZOOM SCOPE BAUSCH & LOMB y posteriormente se digitalizó en AUTOCAD 14.
Para poner en evidencia los cambios geomorfológicos se superpusieron las fotografías arriba mencionadas y una imagen de radar de la bocana (COQUI MAP 183 digital radar mosaic, high resolution, LD west, DF June-jul, 1992) en el STEREO ZOOM para delinear en un mapa los cambios en la línea costera en los últimos cuarenta años.
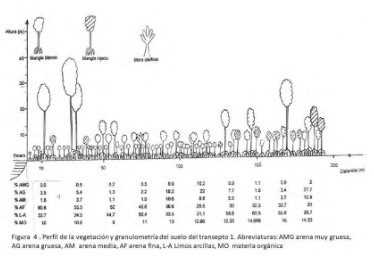
Aspectos edáficos. Las muestras de suelo tomadas para cada parcela se secaron en horno a una temperatura de100 grados Celsius para eliminar el agua contenida; luego se pesaron 100 gramos de cada muestra en una balanza electrónica OHAUS y se procedió a tamizarlos en una batería de tamices. Estas últimas se analizaron por el método de granulometría calculando los parámetros sugeridos por Sánchez (1990): Tamaño promedio de la partícula (Xf), clasificación (Df). De ello resulta una separación granulométrica así: AMG arena muy gruesa, AG arena gruesa, AM arena media, AF arena fina, L-A Limos arcillas, MO materia orgánica (Figura 4).
Manejo de los datos. Para la estructura y zonación (Anexo 1) se partió de los de los datos recolectados en los transectos, en los cuales se evaluaron las características más frecuentemente utilizadas para la descripción de los bosques de manglar tales como: Densidad específica, densidad relativa, frecuencia específica, frecuencia relativa, cobertura específica, cobertura relativa, índice de valor de importancia (IVI). Estas características se establecieron para el conjunto de los individuos y para las categorías diamétricas de brinzal (2.5-5 cm de DAP), latizal (5-15 cm de DAP) y fustal (DAP>15cm) según Sánchez-Páez et al. (1997). La información recolectada se separó en categorías diamétricas para evidenciar más claramente el estado de la vegetación: Estado de regeneración, sucesión y oferta maderera.
Formulas:
-
• Densidad de la especie i: Di =ni/A ni = número de individuos de la especie i, A= área total muestreada.
• Densidad relativa de la especie i: Dri = ni/Ʃn, n= número total de individuos en la parcela.
• Frecuencia de la especie i: fi = ji/k, ji = nú- mero de parcelas que incluyen la especie i, k= número total de parcelas.
• Frecuencia relativa de la especie i: Fri = fi/Ʃf
• Cobertura de la especie i: Ci = ai/A , ai = área del tronco a 1.3 m de altura de i, esta área se calcula a partir de ai = (0.7854)(DAP²).
• Cobertura relativa: Cr = ci/Ʃc
• Índice de valor de importancia: IVIi = Dr+Fr+- Cr
Para saber cómo se distribuyen las
distintas especie de mangle a lo largo de los transectos se estableció para
cada una de ellas el índice de dispersión de Morisita: Formula: Id =
ƩX²-N/N(N-1), n = número de parcelas, N = número total de individuos en todas
las parcelas, X = número de individuos en cada parcela.
A cada uno de los índices obtenidos se le hizo la prueba X² para averiguar si el Id calculado es significativamente distinto de 1,0. Formula: X²= (nƩX²/N)-N, se comparó este X² con el valor en una tabla para n-1 grados de libertad.
También se determinó la probabilidad de hallar un individuo de cada una de las especies de mangle a lo largo de los transectos; para esto se sumó el número de individuos de la especie i presentes en todas las parcelas uno, en todas las parcelas dos, en todas las parcelas tres, etc. Posteriormente se dividió el número obtenido por el total de individuos de todas las especies. Se elaboraron perfiles idealizados de la vegetación de cada uno de los transectos a partir de los datos tomados en cada parcela (Prahl et al. 1990). Estos perfiles guardan la proporción de las especies presentes en cada transecto al igual que las alturas alcanzadas por los individuos.
Resultados
Clasificación
Etnobotánica de Las Especies Maderables del Ecosistema de Manglar. Este resultado se presenta para hacer claridad sobre los nombres
comunes de la especies maderables del manglar utilizadas por las comunidades y
de las cuales se hace referencia en este trabajo. Se encontró que las
comunidades de Virudó y Cuevita identifican nueve especies distintas de mangle,
de las cuales el género Rhizophora tiene
cuatro (mangle blanco, mangle rojo, mangle injerto y mangle “gatión”) (tabla
1). Estas cuatro variedades se van a asumir en el presente trabajo como
morfoespecies por tener características fenotípicas distintas que determinan
diferentes usos. Por esta razón se les separó en la tabla. Más adelante se hace
una descripción más detallada sobre cada una de estas morfoespecies.
Tabla 1.
Clasificación etnobotánica de las especies maderables de mangle
utilizados por las comunidades de Virudó y Cuevita.
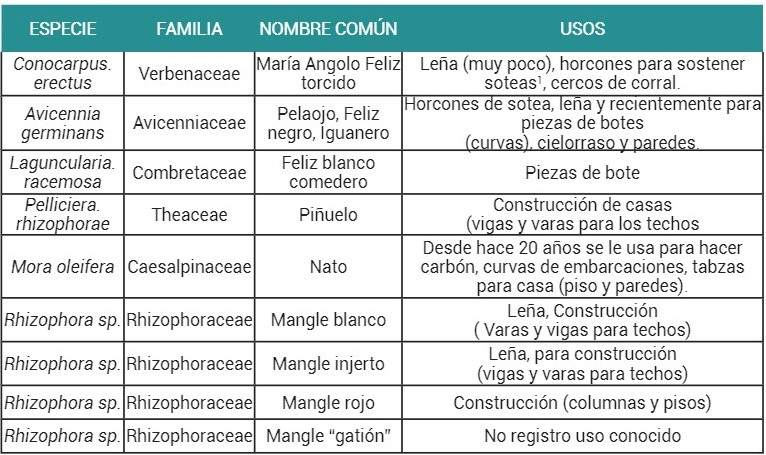 Tabla 1. Clasificación etnobotánica de las especies maderables de mangle
utilizados por las comunidades de Virudó y Cuevita.
Tabla 1. Clasificación etnobotánica de las especies maderables de mangle
utilizados por las comunidades de Virudó y Cuevita.
Etnoecología de algunas especies maderables del manglar. Aquí se hace una aproximación a la ecología de algunas especies maderables del manglar a partir del conocimiento que tienen los nativos de la ensenada de Virudó. Ésta aproximación permite comprender distintos aspectos que determinan el uso de las morfoespecies de mangle y sugieren explicaciones a la zonación mostrada por cada una de ellas.
La figura 2 muestra la zonación ecológica de las diferentes asociaciones vegetales del manglar de Virudó a partir de la fotointerpretación realizada, como son: manglar (dominada por especies del género Rhizophora), natal (dominada por la especie Mora oleífera), piñuelal (dominada por Pelliciera rhizophorae) e impactos antrópicos como la tala raza y formaciones vegetales bajas, estado último que fue estudiado durante esta investigación. Este resultado fue congruente con los datos etnoecológicos aportados por la comunidad en la cartografía social elaborada y en las entrevistas realizadas y por los datos sedimentológicos tomados en las parcelas. A continuación, se describe cada una de las especies que conforman cada una de estas zonas.
Nato (Mora oleifera), Familia Caesalpinaceae: se puede encontrar disperso en el manglar o formando rodales en su parte posterior sobre suelos consolidados (Figura 2). Estos rodales son inundados periódicamente por las mareas más altas pero con bajas concentraciones de sal. El nato florece y muda el follaje en el mes de mayo, en este tiempo llega un gusano a comer sus retoños. En octubre bota los embriones coincidiendo con las pujas más altas del año. Después de cortado puede retoñar siempre cuando haya sido con luna menguante. “De treinta natos que se encuentren aproximadamente la mitad están huecos”. Estos huecos son aprovechados por distintas especies para hacer sus nidos. Entre ellas el lobo, el cuzumbí, el oso hormiguero y distintos pájaros. Las abejas también acostumbran a construir panales en esos huecos.
Piñuelo (P. rhizophorae) Familia Theaceae: abunda en las partes firmes, generalmente en la zona de transición de manglar a natal. Se puede encontrar mezclado con mangle o con nato o formando rodales. Todo el año produce embriones. El crecimiento es más acelerado que el del Nato y su madera es poco resistente (Figura 2).
Mangle (Rhizophora spp.) Familia Rhizophoraceae: el caso de éste género es muy interesante. En la zona norte del Bajo Baudó (Chocó), los nativos lo separan en cuatro morfoespecies: Mangle blanco, Mangle rojo, Mangle injerto y Mangle gatión. La diferenciación se hace con base en su zonación en el manglar, la forma de sus raíces, la textura y grosor de la corteza, su estructura, forma de crecimiento y las características de su madera (figura 2).
Mangle “gatión”: generalmente se le encuentra en las orillas de los esteros, aunque en raras ocasiones se le puede hallar manglar adentro en suelos muy inestables. Este árbol no forma fuste, sino que va creciendo para los lados (crecimiento plagiotrópico), “como gatiando[2] ” por eso su nombre “gatión”, y no alcanza alturas superiores a los 10 metros. Durante todo el año produce embriones, aunque entre abril y mayo encuentra su mayor producción. Su madera es blanca y difícil de rajar. De las cuarto morfoespecies es la que posee las hojas más anchas.
Mangle blanco: comparado con el mangle injerto y el rojo, se le encuentra en las partes más inestables del manglar colindando con el gatión; sus raíces son más altas, su crecimiento es más acelerado, alcanza mayores alturas (30 y 45 m) y el fuste es recto. Su corteza es más delgada, lisa y de color grisáceo. Su madera es completamente blanca, se utiliza mucho para leña por ser fácil para rajar y su humo no irrita los ojos. La hoja es delgada.
Mangle injerto: con respecto al blanco, sus raíces son más bajas y se ubica en suelos más consolidados entre el mangle rojo y el blanco. También alcanza altas tallas (30 y 40 m). Su corteza es más oscura, rugosa y fragmentada en anillos. Cortando transversalmente el tronco se observa el corazón o duramen de color rojo. Las hojas al igual que las del mangle blanco son delgadas. Su madera es más resistente que la del blanco y es utilizada para construcción. También se usa para leña pero menos que el blanco, es más difícil de rajar y su humo irrita los ojos.
Mangle rojo: se le encuentra más fácilmente en las partes más firmes del manglar, cerca de los natales. Sus raíces son “sentadas”, más bajas que las del blanco y el injerto. No alcanza la misma altura de estos (30 m) y su crecimiento es marcadamente lento. Generalmente el fuste es irregular. Su corteza es rojiza, más gruesa, más rugosa y con anillos y entrenudos menos distanciados que los del injerto. El corazón rojo o duramen del tronco es más grande que la del injerto, solo tiene máximo 3 cm de madera blanca o albura hacia el borde. Sus hojas son anchas y redondas, parecidas a las del gatión y persisten en el tallo. Es decir que las hojas viejas se secan en el tallo sin desprenderse rodeando las nuevas. Su madera es la más dura y difícil de astillar. Se le utiliza mucho en construcción por su gran resistencia.
Historia de Uso y Explotación del Ecosistema de Manglar. La historia de la costa pacífico colombiana se inscribe dentro de la historia general de explotación de los recursos naturales del país. Explotación agenciada en muchos casos por el estado, mediante el otorgamiento de licencias de explotación a particulares o por consentimiento tácito, y que se ha hecho en zonas rurales pobres dónde sus habitantes, carentes de título sobre sus tierras, solo han participado como mano de obra asalariada en las labores de extracción. Muy comunes son las menciones que se hacen en las narraciones orales de los pobladores a estas explotaciones y que se remontan al siglo pasado. Ellos se refieren a épocas de explotación intensiva de diversos productos: época de la tagua, época del caucho, época del banano, de las pieles de los animales, de la cortecera y de la pilotera. Cabe resaltar que para todas estas épocas, eran personas foráneas de Panamá o del interior, las que venían a comprar estos recursos a los nativos.
Como lo anota Friedmann (1989) estas comunidades históricamente han vivido un proceso caracterizado por frecuentes e intensos cambios en la fisonomía económica, política y social de la región por la intervención de agentes externos, nacionales e internacionales interesados en la explotación de sus recursos. La explotación del ecosistema de manglar, que es el tema del presente trabajo, se enmarca dentro de esta misma lógica de economía extractiva.
Las comunidades negras e indígenas tradicionalmente han aprovechado la madera del mangle para la construcción de embarcaciones, instrumentos de trabajo, viviendas, combustible; pero es a partir de la demanda en las ciudades del interior que se rompe esta actividad artesanal por una economía de extracción comercial con la llegada en 1954 de la empresa Lizcano Hermanos a la parte norte del Municipio del Bajo Baudó. Esta empresa denominada por las comunidades como “la cortecera” hacía parte de la industria de curtiembres, una de las más importantes del país para esa época. Estaba cimentada principalmente en la acción fijadora que, sobre las proteínas de la piel animal, ejerce el tanino, sustancia extraída de la corteza del mangle. (Rhizophora spp.). Para abordar la historia de explotación y uso del manglar del área de estudio es necesario delimitar tres momentos: Antes de la llegada de la empresa Lizcano Hnos, durante el periodo de explotación de la empresa y después de que se fue la empresa. Estas separaciones se hacen para comprender de forma clara las relaciones existentes entre las distintas dinámicas de extracción y los cambios acontecidos en el ecosistema y en la cultura.
Historia del Manejo antes de “la cortecera” Usos. Antes de la llegada de la empresa Lizcano hnos, el uso maderero del manglar estaba restringido a madera para leña y a la construcción de casas, de una manera muy limitada, pues las dimensiones de la madera requerida para estas necesidades no se hallaba con facilidad. Esto se debía a la preponderancia de árboles en el manglar con diámetros superiores (en promedio 0.75 metros) a los requeridos. Para encontrar un pilote (como se les llama a los árboles de mangle de 0.40 metros de diámetro) había que ir lejos, a zonas donde se le podía encontrar más fácilmente (p.e. esterones de Virica). Por esta situación gran parte de la leña utilizada provenía de la montaña (Jaguillo, guayabo y carbonero).
La madera de mangle (Rhizophora spp. y P.rhizophorae) que se utilizaba para la construcción de viviendas era la que tenía tamaño de viga (0.10 metros de DAP), vara (0.8 metros de DAP) y pilote. Las dos primeras se utilizaban para colgados y enrazaduras de los techos y el pilote para cimiento de la vivienda. El piso era de palma, fuera barrigona (Iriartea ventricosa), memé (Wettinia quinaria) o zancona (Socratea exorrhiza); las paredes de guadua y el techo de hoja de palma. Este tipo de construcción cambiaría radicalmente con la llegada del primer aserrío, y la mecanización del corte de madera que daría la posibilidad de usar madera aserrada en forma de tabla, remplazando así el uso de la palma.
Como leña se prefería y se sigue prefiriendo el mangle injerto y el mangle blanco (Rhizophora spp.), por su facilidad para astillar y porque su humo que no arde a los ojos. Con un árbol que se tumbaba alcanzaba para el consumo doméstico de una familia por casi dos meses.
Descripción del ecosistema de manglar antes de “la cortecera” La siguiente descripción se hizo con base en los testimonios de los entrevistados, y muchas de sus apreciaciones se hicieron comparándolas con la actualidad.
En los manglares antes de la cortecera “abundaba mucho animal”, mucho más de lo que se encuentra hoy en día. Se conseguían muy fácilmente vivalvos como la piangua (Anadara tuberculosa) y el ostión (Crassostrea columbiensis), cangrejos como el meón y el halacho (Ucides cordatus), caracoles como piacuil (Littotina zebra) y el puyudo (Thaïs sp.), iguanas (Iguana iguana), lobo (Procyon cancrivorus), aves y peces en general. En una hora se podía recolectar hasta una “lata[3]” de piangua, hoy esto mismo se consigue buscando durante 6 horas.
El manglar estaba dominado casi completamente por las distintas morfoespecies de Rhizophora (mangle blanco, injerto, rojo y gatión), de estas el mangle rojo abundaba mucho más de lo que abunda hoy. Para poder encontrar un piñuelo (P. rhyzophorae) había que recorrer hasta 200 metros. El piñuelo se encontraba preferiblemente en la zona de transición de manglar a natal, fuera mezclado con mangle rojo o con nato. En la zona de los natales se veían algunos árboles de montaña como el machare (Symphonia globulifera), otobo, suela (Pterocarpus officinalis), además de naidí (Euterpe sp.).
En cuanto al DAP de los árboles, se encontraban palos entre 0.50 y 0.75 metros. “El manglar era surtido de mangles gruesos”, “en 100 metros de trayecto se encontraban hasta 50 palos grandes de 40 pulgadas”, “ los mangles que había eran de abarcadura (que no se al- canzaban a abarcar con los brazos)”. Se encontraban de estos diámetros (entre 0.50 y 0.75 metros de DAP) tanto en las orillas como en las partes internas. No se encontraban fácilmente mangles de diámetro de vara y viga (0.08 y 0.10 metros de diámetro). De los pequeños los que más abundaban tenían un promedio de 0.30 metros.
as raíces de los mangles alcanzaban alturas entre 3 y 4 metros, y era tal la densidad de ellas que se dificultaba mucho caminar. Los manglares eran muy sombreados debido a la densidad del dosel. Esta situación dificultaba el crecimiento de embriones y juveniles. Las raíces o “ñanglas” estaban completamente invadidas por algas y ostiones y los troncos por “chupayas ”(bromelias).
La bocana con todo su sistema de esteros era significativamente más profunda que lo que hoy es, las playas no se inundaban y los poblados que estaban en sus orillas no vivían el proceso acelerado de erosión, que hoy están viviendo.
Dinámica Estructural y Ecológica durante “la cortecera” Con la llegada de la empresa Lizcano Hermanos a la zona norte del municipio del Bajo Baudó en 1954. Manuel Rentería Gómez, proveniente de Ijuá[4], llegó a Pilizá[5] como contratista de la empresa. En Pilizá fue donde se instaló el primer centro de acopio de corteza de mangle. Con “Cancharina”, como le llamaban a Manuel Rentería llegaron ocho hombres expertos en extraer la corteza para que le enseñaran a los nativos el oficio. Al cabo de un año, de Pilizá se amplió la extracción al sector de Purricha[6], para finalmente llegar a la bocana de Virudó en 1958. Cuando la empresa llega, las actividades principales de los nativos eran la pesca, la cacería y la agricultura. Sin embargo como fueron aprendiendo el oficio, hombres mayores y jóvenes ayudados por sus familias, dedicaron más tiempo a ésta actividad extractiva. Aproximadamente el 90 % de la población masculina adulta se vinculó a este trabajo.
Según los habitantes de la región, la empresa trajo muchos beneficios a la comunidad. La salud y la educación mejoraron. El transporte, utilizado para llevar a Buenaventura la corteza, era aprovechado para transportar los enfermos. Se nombró profesores pagados por la empresa. Se construyeron puestos de salud y escuelas en los poblados más grandes. Los costos médicos de personas accidentadas o enfermas, que trabajaban en la cortecera, eran sufragados por la compañía. Y en diciembre eran pocos los que no recibían regalos. El dinero comenzó a circular, atrayendo mucho el comercio. En el poblado de Pavasa se instaló la discoteca y la tienda “más importante de la zona norte del municipio”, cuyo dueño era el hijo de “Cancharina”. A éste lugar llegaban trabajadores de todo el sector a beber y a comprar víveres con lo ganado en el trabajo de extracción. Era usual que los trabajadores fiaran estos productos mientras llegaba el pago.
Muchas familias disminuyeron su actividad agrícola por dedicarse a extraer corteza. El alcalde del municipio se vio obligado a sacar un decreto que exigía a cada familia sembrar 100 matas de plátano. La escasez de alimentos había incrementado dramáticamente el robo de las cosechas.
En la población es unánime la idea que éste periodo de la cortecera fue beneficioso para la economía de la región, “trajo progreso”.
Formas de explotación. La actividad de cortecear (acto de quitar la corteza) era una actividad familiar. En ella participaba de alguna forma toda la familia. El padre, era quien tumbaba el árbol y le quitaba la corteza, los hijos menores y a veces las mujeres la recogían y la embarcaban en la canoa. El papel de la mujer era fundamentalmente la culinaria, ella se levantaba temprano a preparar los alimentos o merienda que el marido y los hijos se llevaban. Los hombres a su vez trabajaban en cuadrillas de 3, 4 o 5 personas. Escogían el lugar donde iban a extraer y se colaboraban.
Recién llegada la empresa Lizcano, la actividad extractiva se hacía en los lugares aledaños a los puestos de acopio que se ubicaron en un principio en los poblados o cerca de ellos. Los primeros “palos” que se derribaron eran los más cercanos a estos puestos y que estaban en la orilla. En la medida en que se iban escaseando en estos lugares, la actividad se hacía cada vez más adentro en el manglar y más lejos de los puestos. Para poder sacar la corteza de la parte interior del manglar se hacían “picas”, o sea canales perpendiculares a la orilla por donde podían pasar embarcaciones en marea alta. De esta forma se creó una compleja red de canales que atravesaban los manglares de estero a estero.
Los árboles que se utilizaban para descortezar eran el mangle blanco, mangle injerto y mangle rojo, todos morfoespecies del género Rhizophora. El más apetecido era el rojo, porque era el de mayor grosor en la corteza, y por lo tanto el que más rendía económicamente.” Más peso, más plata”. La escogencia del palo a tumbar dependía de su tamaño y de la facilidad para ser descortezado. Para esto se cortaba parte de la corteza y se probaba su adherencia al tronco. Muchos árboles que se derribaron se perdieron por esta razón, es decir que una vez tumbados era imposible extraerles la corteza. Para tumbar un palo, primero se observaba si tenía algún grado de inclinación para así escoger qué raíces cortar y facilitar su caída. Para cortar las raíces se usaba el hacha. Demorándose de una a tres horas cortándolas hasta que cayera el árbol.
Cuando recién comenzó la extracción, esta labor se dificultaba mucho. La gran densidad de árboles y raíces no daba casi espacio para trabajar. Como los árboles escogidos eran los más grandes (0.75 metros de DAP), al caer se llevaban consigo a los más pequeños. Con tumbar un solo palo podían caer hasta tres palos más, abriendo un claro considerable en el dosel. Una vez en el suelo, el árbol comenzaba a ser descortezado. Primero la persona se paraba encima del tronco y comenzaba a cortar la corteza longitudinalmente ayudándose con el hacha. Esto se hacía tanto a la derecha como a la izquierda del lugar donde estaban los pies del cortecero. Después con la misma hacha se despegaba la corteza. Cuando se dificultaba su desprendimiento se le golpeaba con un palo para que aflojara. Si el tronco del árbol había quedado muy enterrado en el fango, la parte de corteza que quedaba hundida se perdía.
De un solo palo se podía extraer hasta dos toneladas de corteza, peso que fue disminuyendo en la medida que avanzaba la explotación. La corteza desprendida se cargaba en la o las canoas y se transportaba, hasta el puesto de acopio donde se pesaba y se pagaba. En un comienzo se pagaban 10 pesos por una tonelada y antes que la compañía partiera se pagó a 35 pesos.
En esta actividad eran muy frecuentes los accidentes, en ocasiones mortales. Las personas mayores cuentan que compañeros suyos murieron aplastados o quedaron lisiados al caerles encima el árbol derribado. Las marcas, fueran heridas de hacha o causadas al caer los árboles, les recuerda esta época de sus vidas. Además de estos accidentes, ellos hablan de la gran cantidad de riesgos naturales que corrían en los manglares. Los alacranes, las arañas, las culebras y los peces sapo; eran una continua molestia.
Los nativos del lugar solo sacaban corteza durante 15 días al mes en el periodo de quiebra[7] de la marea, el resto del tiempo lo usaban para pescar y cuidar la finca. A diferencia de los nativos los hombres que vinieron del sur de la costa chocoana, traídos por los contratistas (40 en total), dedicaban todo el tiempo a esta labor. Trabajaban incluso por las noches cuando había luna llena. Cocinaban en el mismo manglar. Recogían las conchas de piangua que encontraban por montones y las cocinaban junto al arroz y al plátano que llevaban. Para estos hombres y sus familias se construyeron dos poblados, ubicados cerca de los principales lugares de explotación. Uno en el estero El Encanto y otro en Morro Blanco.
Zonas de extracción. Como se mencionó arriba, los primeros sitios donde se extrajo corteza fueron los más cercamos a los poblados o centros de acopio. En el poblado de Virudó se comenzó en el estero bocón y en el río Virudó. En Cuevita se comenzó en Morro Blanco. Una vez se fueron agotando estos sitios, los que vivían en Virudó comenzaron trabajar los ríos de Pital, Pavasa y el estero Peñitas. Los Cueviteños pasaron al Encanto y a Caimanero.
En cada uno de estos lugares, la dinámica de explotación fue la misma. Se comenzaba en las orillas y se terminaba hacia adentro del manglar. Aunque según lo expresado por los entrevistados, la dificultad de trabajar al interior impedía que se explotara con la misma intensidad que en la orilla. De la misma forma los lugares de extracción iban cambiando, primero se ubicaban cerca de la boca del estero y finalmente río arriba hasta dónde llegara el manglar. Cuando ya no se hallaban árboles grandes río arriba, la dinámica volvía a empezar, pero ahora utilizando los árboles más pequeños que habían quedado. “No hubo sitio que no fuera tocado por la cortecera”. En Virudó se recuerda que el último sitio explotado fue en el estero Bocón y en Cuevita en el Encanto.
La ubicación de los centros de acopio de corteza fue acorde con la dinámica de explotación. Es decir que los puestos se iban trasladando cerca de las nuevas áreas explotadas. En Virudó el primer puesto se construyó frente al poblado en la Isla de Barrial (Figura 2), luego cerca de la boca del estero Bocón y posteriormente en el sector de Peñitas. Finalmente, el puesto se instaló nuevamente cerca de la boca del estero Bocón. Según los entrevistados fue en éste último estero donde más se concentró la actividad extractiva, seguido por el río Virudó y Peñitas. En Cuevita, el primer puesto estuvo en Morro Blanco y finalmente en el Encanto. Estos fueron los sitios más explotados.
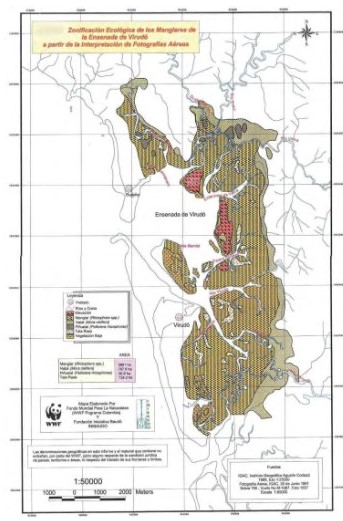 Figura 2.
Zonificación ecológica de los manglares de la ensenada de Virudó
Figura 2.
Zonificación ecológica de los manglares de la ensenada de Virudó
Volúmenes de explotación. Para tener una idea de los volúmenes de explotación es importante tener en cuenta la cantidad de población que se dedicó a la cortecera. Según una fotografía aérea, para el año de 1961 el número de casas para Cuevita y Virudó era de 40 y 61 respectivamente. Aquí se discriminó entre volúmenes de extracción por persona y el total para la bocana.
Según las personas de mayor edad entrevistadas que participaron de la explotación en sus inicios, los árboles de mangle más grandes podían aportar entre 2 y 3 toneladas de corteza. En promedio los árboles derribados daban entre una y una y media toneladas. Este promedio era el mismo para la explotación diaria y por persona. Los hombres provenientes del sur, los más prácticos, sacaban hasta tres toneladas después de haber tumbado igual número de árboles. En la bocana un mínimo de 100 personas se dedicó a esta labor extractiva.
Según Manuelito Rentería, contratista de la empresa, el espesor de la corteza de un árbol podía alcanzar cinco centímetros, que aportaban tonelada y media. Al final de la cortecera y después de un estudio que hicieron unos ingenieros contratados por la empresa, el espesor promedio de la corteza era de un centímetro y medio. Lo anterior coincide con la información aportada por los entrevistados más jóvenes. Estos que se vincularon en la etapa final de la explotación, afirman que el promedio de explotación oscilaba entre media tonelada y una tonelada. Y para lograr estos valores había que derribar hasta cuatro palos. Como lo afirma Adolfo Asprilla (46 años de edad) quien se vinculó a la cortecera en 1968: “se sacaba entre 700 y 800 kilos en un día habiendo tumbado cuatro palos”.
Para transportar la corteza a Buenaventura la empresa utilizaba tres barcos, el mayor de los cuáles tenía una capacidad de ochenta toneladas (Delgado & Flórez 1965). Sin embargo, si la producción lo ameritaba, entraban barcazas con capacidad para 350 toneladas (afirmación de Manuelito Renteria). Según el contratista de la empresa, “de Pilizá, salía un barco cada 8 días, había veces que entraban hasta dos barcos al tiempo. El Balboa, el Micay y el Utría eran los barcos de la compañía. Otra forma de sacar corteza era en pangones[8]. Al mes podían salir hasta 500 toneladas en toda la región (de Cuevita a Virudó). A veces no se alcanzaba a sacar la producción. El Balboa hacía hasta tres viajes repetidos, se alcanzaba a dañar la corteza porque se secaba demasiado,”.
En el resto de las entrevistas se plantea que a comienzos de la cortecera, de la bocana de Virudó salían dos barcos mensualmente, a veces entraban más embarcaciones entre pangones y barcazas. Las embarcaciones variaban en sus capacidades, oscilaban entre 50 y 120 toneladas. Es decir que de la bocana salía un mínimo de 100 toneladas al mes. Ya en el periodo final de la cortecera, desde 1965 aproximadamente, a la bocana sólo entraba un barco cada dos meses. Según Delgado y Florez (1965), en 1965 de toda la región (desde el río Pilizá hasta Virudó) salían 250 toneladas de corteza al mes.
Fin de “la cortecera” Cuando el estado de los bosques de manglar no respondió a las necesidades de explotación, la industria Lizcano Hermanos se marchó. Esto sucedió a finales de la década del setenta. El puesto de donde salió el último embarque quedaba cerca de la boca del estero Bocón.
• Descripción
del ecosistema: Para esta descripción se va a ser fieles a las impresiones de los entrevistados. Ellos, con sus ojos, vieron como quedó el ecosistema y que pasó con él una vez que la “cortecera” se marchó.
“Los manglares quedaron desolados”, ”con la
cortecera se acabó todo”, ”quedaron como una socola”, “después de la cortecera
los mangles quedaron asolados”, “los manglares quedaron destruidos, se secaron9
los esteros”. ”Cuando la cortecera se
fue quedaron poquitos árboles grandes que no se podían descortecear, quedaron
limpios los manglares, entraba mucho el sol y los cangrejos se fueron”. “Con
el sol se comenzaron a morir la piangua, el alacho y el piacuil bastante, también el
pulludo. Con Una entrevista que recoge las distintas impresiones de cómo quedó
el ecosistema fue la realizada a Manuel Antonio Córdoba, profesor jubilado y
antiguo proveedor de la empresa Lizcano Hnos. Manuel cuenta que después de la
explotación todo era “desolación”, ”la tala del mangle trajo como consecuencia
la erosión de los manglares, porque los árboles cubrían el suelo y al
desnudarlo el agua caía directamente sobre el barro. El peso del árbol y las
raíces detiene la erosión, entonces sin árboles se da el derrumbe de los
manglares y también el relleno de la bocana. Se fue la jaiba, el camarón, el
cangrejo, la piangua y el piacuil. Se rellenaron los esteros, esta bocana era
profunda pero entonces todo ese barro se iba al estero, entonces se iba
llenando, se iba llenando. El pescado se ahuyentó, porque en las raíces de los
mangles había lo que se llama limo, una especie de alga, eso lo comía mucho el
pez, pero como el mangle tiene ese tanino y los peces respiran por branquias,
les endurecía las agallas y no se facilitaba la respiración, por eso huía. Al
final de la cortecera árboles pa´cortecear se veían pero muy escasos, eran
pequeños y se encontraban muy adentro”.
“A pesar de toda esa destrucción, en el año de 1975 los árboles grandes que quedaron en pie dieron una producción de embriones como nunca se había visto. El mangle se comenzó a reforestar rápidamente. “No dio tregua, no dejó que otra especie ocupara su lugar”. “Una cosa que tiene el mangle es que él se reforesta muy rápido”.
Momento después de “la cortecera” y Comienzo de la época de “la pilotera” Cuando la industria de la corteza de mangle se acabó, pasaron entre dos y tres años antes que llegara la nueva época de extracción, “la pilotera”. La época de “la pilotera” se refiere al periodo en el cual se extrajo del manglar madera de mangle (Rhizophora spp.) con un DAP aproximado de 40 centímetros. A este tipo de madera se le denomina “pilote”. Según refieren los entrevistados, esta madera era usada para postes de luz en las ciudades y para construcción. De Buenaventura, donde se almacenaba la producción de toda la costa, se exportaba a Estados Unidos. Es importante anotar que sobre este periodo de explotación del ecosistema de manglar poco o nada se ha escrito. En varios textos se alude a lo que fue la explotación de la corteza, pero pocos hablan sobre lo que pasó con “la pilotera”.
En la bocana de Virudó la extracción de pilote comenzó con la llegada de los señores Luis Mario Garrido y Luis Alirio Muñoz. Provenían de las bocas del San Juan donde tenían un aserrío en Isla Monos. A su llegada montaron un aserrío en donde estuvo el último puesto de acopio de la cortecera, este sirvió para almacenar los pilotes. Los primeros pilotes que se cortaron eran de Piñuelo (P. Rhizophorae), pero su madera no respondió a las necesidades de resistencia. Se rajaba fácilmente. También se probó con Iguanero (A. germinans) y tampoco sirvió. Muchos fueron los árboles que se cortaron y no sirvieron. Finalmente la explotación se concentró en el mangle rojo, blanco e injerto (Rhizophora spp.). De estos se prefería el rojo, por su mayor resistencia.
Se entrevistó la actividad de explotación y se encontró que habitantes locales (en la investigación identificados como Muñoz y Garrido) estuvieron sacando pilote a Buenaventura durante un año, después se dedicaron exclusivamente al aserrío. Luego, en octubre de 1983 llegó otro empresario (en el estudio Eduardo Torres) a incentivar nuevamente la corta del pilote. Con este señor comenzaron a llegar otros contratistas, algunos con permiso del Inderena y otros sin él. Algunos nativos se vincularon también de contratistas. Así como la época de la cortecera, este nuevo periodo fue de bonanza económica. El dinero volvió a circular. Los hombres, en su mayoría jóvenes y viejos corteceros, se dedicaron a esta nueva actividad. Todos lo dicen: “la época de la pilotera fue más dura que la cortecera”.
Los manglares que ya no ofrecían árboles para descortezar estaban surtidos de pilote gracias a una rápida regeneración. Estos eran los que comenzaron a crecer cuando tumbaron los primeros árboles durante la cortecera.
Formas de explotación. El proceso para cortar un pilote era más sencillo que el utilizado en la cortecera. Se usaba igualmente el hacha pero el esfuerzo era menor. Los árboles ahora eran más pequeños. Se preferían aquellos cuyo fuste era recto se cortaban sin distinguir entre mangle blanco, rojo o injerto. Una vez en el suelo se le quitaba la corteza y se le cortaban todas las ramas. Luego se le transportaba en hombro hasta la orilla donde los iban apilando.
Para sacar los palos que se tumbaban adentro en el manglar se aprovechaban pequeños esteros auxiliares o se hacían picas de dos metros de ancho. Cada pica tenía sus dueños. Los que las hacían eran los únicos que tenían derecho a usarla. Una vez terminada la pica, el grupo se dividía en parejas por la zona aledaña para tumbar, pelar y transportar los árboles de servicio. En esta época fue mayor el número de picas que se hicieron. Evidencia de ello son las herraduras que hoy utilizan los habitantes de la región para acortar camino rumbo a su finca. La corta y el transporte del pilote se hacía en dos tiempos. Se cortaba durante el periodo de quiebra[9] y se sacaba con la puja11 hasta el sitio donde iba a ser recogido por el barco. A esta actividad dedicaban hasta tres semanas al mes, la semana restante era utilizada para subir a la finca a buscar comida. El área aledaña a la pica o al estero auxiliar se agotaba después de tres meses aproximadamente.
Cuando en la región había una cosecha importante, como el arroz, la explotación se paraba casi por completo durante un mes.
Zonas de explotación. La dinámica de explotación del pilote de mangle se dió de la misma forma a como ocurrió con la cortecera. Primero se comenzó a explotar en los alrededores del primer puesto de acopio que quedaba cerca al poblado. Así fue para Cuevita y Virudó.
Las zonas de explotación se fueron desplazando hacia arriba de los esteros y lejos de los poblados en la medida en que se iba agotando el recurso cerca de los poblados. En Virudó los primeros pilotes se sacaron frente al aserrío actual (Figura 2) y en Cuevita del estero El Encanto. Los puestos de acopio se fueron trasladando a las nuevas zonas de extracción. Los hubo hasta donde el barco pudiera entrar a cargarlos.
Los esteros de dónde se extrajo Pilote fueron (anexo A): Virudó, Bocón, Pavasa, Pital, Peñitas, Esterones de Viríca, Morro Blanco, El Encanto, Caimanero, Clementina, Viríca y Jella. De estos, los sectores de Morro Blanco, El encanto y Bocón fueron los más explotados. Pero al igual que en la cortecera “no hubo zona que no fuera tocada”.
Volúmenes de explotación. Fueron pocos los hombres mayores y jóvenes, que no dedicaron parte de su tiempo a sacar pilote. Aproximadamente un 90 % de ellos trabajó en “La Pilotera”. En 1990 Cuevita contaba con 47 casas y 191 habitantes, y Virudó 145 casas y 667 habitantes (datos SEM). Por lo menos 170 hombres sacaban pilote.
En los primeros años de “la pilotera” los hombres sacaban entre 15 y 20 pilotes en un día. Esta cantidad dependía de la habilidad que tuvieran. En promedio, como se mencionó anteriormente, ellos dedicaban a este oficio 3 semanas al mes. Sin embargo no trabajaban de forma continua, durante estas semanas. La labor de tumbar, pelar y sacar un pilote era muy extenuante. Lo anterior hacía que en promedio una pareja sacara al mes entre 150 y 225 pilotes. En esta etapa inicial, a la bocana entraban por lo menos dos embarcaciones al mes. Cada una de ellas podía transportar entre 1500 y 2000 pilotes. Aunque en el periodo de mayor auge, cuando “llegaban muchos contratistas”, entraban hasta diez embarcaciones. Ya a finales de esta época, antes de 1993 (año en el que se establece la veda por parte del Inderena), solo entraba un barco cada dos meses. Obviamente debido a la progresiva escasez del recurso por la exagerada intensidad con que se dió esta explotación.
A pesar que en 1993 se establece la veda del mangle, la explotación no termina allí. En la época de “la pilotera”, paralelamente pero en menor intensidad, se estaban sacando varas y vigas de mangle (con 0.07 y 0.10 metros de DAP respectivamente). La extracción de mangle con estas dimensiones se intensificó una vez terminó “la pilotera” y a pesar de la prohibición. De la bocana salieron 5000 palos (entre varas y vigas) al mes hasta el año de 1998. Entre este año y el año 2000, un nativo de la región ha sacó no más de 12 viajes de 1000 palos cada uno. La disminución se ha debido a la toma de conciencia de los pobladores y a los tardíos controles de CODECHOCO (Corporación Autónoma del Chocó).
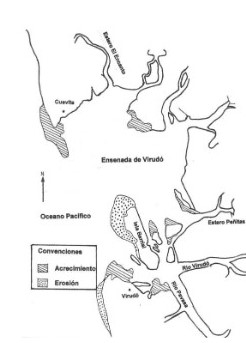
Reconstrucción Histórica de la Dinámica Costera de La Bocana. La siguiente reconstrucción histórica es el resultado de las entrevistas. En ella se integran los cambios vistos por los entrevistados, los cambios narrados por sus padres y los análisis que ellos mismos hacen sobre cómo ha sido la dinámica costera. La forma que tiene la bocana hoy es muy distinta a como era antes. Los mayores cuentan que en la época de sus padres hace más de cien años”, el morro de Civira estaba conectado con la isla de Barrial (figura 3). “Todo eso era una sola isla”, Civira estaba rodeada de bosque y había casas que quedaban hacia fuera del morro, mirando al mar. “El mar no llegaba a Civira”. El poblado de Cuevita quedaba muy cerca de la isla de Barrial, tanto que uno podía gritar hacia el otro lado para que lo pasaran en canoa. En ese entonces la bocana era mucho más estrecha y la entrada de las embarcaciones estaba enfrente de Cuevita.
La parte de Cuevita que daba hacia la bocana era un barranco. Este comenzó a derrumbarse por la alta actividad sísmica. “En esa época temblaba bastante y cuando venía el temblor los pedazos de barranco se iban cayendo y en estico la marea ya estaba alta, llegaba con maremoto, fueron tres años seguidos”. Ya en las primeras décadas del siglo XX, desde Barrial no se alcanzaba a escuchar cuando alguien gritaba desde Cuevita. Civira había quedado como un accidente geográfico separado de la isla Barrial. El mar había lavado la tierra firme que lo rodeaba, pero un banco de arena, que no anegaba lo unía con la isla de barrera de Virudó. La cabecera de la inspección de Virudó en ese entonces quedaba más cerca de la isla de Barrial de lo que hoy está. Virudó tenía una sola caleta por donde entraban las embarcaciones y estaba protegido de la arremetida de la marea por un cinturón de mangle.
En 1950 ya “se había ido mucha tierra agua abajo”, Cuevita lentamente seguía retrocediendo por la erosión. Sin embargo “un higuerón grandísimo” la protegía aún. Pero lo tumbaron y el agua siguió lavando el terreno hasta derribar las casas. En ese momento la isla de Barrial seguía retrocediendo también. El manglar que enfrentaba la arremetida del mar comenzaba a secarse.
La bocana, que aún era profunda comenzó a rellenarse con la llegada
de “la cortecera”. El barro, en los manglares desolados, quedó a merced de la
acción del agua y fue a dar a los esteros y a la bocana. Las playas comenzaron
a ser inundadas por las mareas grandes y el morro de Civira se desconectaba
periódicamente con cualquier marea.
En Virudó tumbaron la franja de manglar que lo protegía y el barrio “Las Brisas”, que quedaba frente a Barrial se comenzó a ir con el agua y Virudó tuvo que trasladarse hacia adentro. Muchas casas fueron derribadas y reconstruidas. “En el año de 1975 las últimas casas en dirección opuesta a la orilla estaban donde hoy quedan las primeras, el agua se ha llevado más de cien metros de terreno”. Cuevita mientras tanto también siguió cediendo y trasladándose. “Cuevita estaba donde hoy está el palo de pelaojo”. Este árbol está hoy a doscientos metros de donde está Cuevita y se aniega completamente con marea alta.
En estos momentos el proceso de erosión continúa. Tanto Virudó como Cuevita se inundan completamente con las mareas más grandes del año. Esto ha obligado al traslado de Cuevita doscientos metros monte adentro. Virudó, donde se puede navegar por las calles con estas mareas, tiene el proyecto de ser trasladado también. La isla de Barrial está más reducida, la arremetida del mar continúa secando el manglar y amenaza con llegar donde hoy están las casas. El manglar se ha secado “por la arena como este busca el barro y lo que se está depositando allí es arena y la ola golpea y se lleva el barro”. “Hoy para ir de Cuevita a Virudó a canalete se gasta uno una hora más de lo que se gastaba hace 50 años”. Un ejemplo de la dinámica sedimentológica del manglar se ilustra en la figura 4, que muestra como estas corresponde a la estructura misma del ecosistema.
 Figura
3.
Mapa de los cambios en la línea de costa en la
ensenada de Virudó entre los años 1961 y 1992.
Figura
3.
Mapa de los cambios en la línea de costa en la
ensenada de Virudó entre los años 1961 y 1992.
Discusión. Es claro, después de ver cuál ha sido la historia de uso y explotación del ecosistema de manglar de la ensenada de Virudó y Cuevita, que este ecosistema desde 1954 ha sido sometido a una fuerte tensión motivada por las actividades antrópicas ejercidas sobre él. Tensión que en las últimas décadas ha disminuido sin detenerse por completo, como se puede observar en la figura 2 la cual muestra fotografías de zonas con tala raza.
La explotación del manglar se concentró en una primera instancia en la extracción de la corteza del mangle (Rhizophora spp.) como fuente de Taninos para la industria del Cuero y posteriormente en la extracción de madera, utilizada en la industria de la construcción y como postes de luz. Estas actividades han afectado de forma marcada la dinámica general del ecosistema de manglar.
Estructura del ecosistema del manglar. El anexo 1 ilustra aspectos descriptivos de la estructura del manglar de Virudó en varios esteros representativos. Comparando el estado del ecosistema de manglar antes de que llegara la empresa Lizcano hermanos, cuando dominaban árboles de mangle (Rhizophora spp.) con DAP´s superiores a 0.5 metros, con el estado actual donde dominan las categorías diamétrica de Brinzal y Fustal; es evidente que ha habido una disminución en su complejidad estructural (altura y DAP). Antes de la época de “la cortecera” los árboles de mangle no tenían mayor uso. Los diámetros más abundantes eran demasiado grandes para poder ser utilizados con las herramientas que poseían los nativos en ese entonces. Hoy para encontrar un árbol suficientemente grande para extraerle la corteza hay que recorrer más de cien metros en el manglar.
Muy diciente es el orden en el que se sucedieron las épocas de explotación. Comenzó por la corteza cuando abundaban árboles que aportaban grandes volúmenes de corteza, siguió con el pilote (DAP igual a 40 cm) y terminó con la extracción de la vara (7 cm de DAP). Cada una de estas épocas coincidió con los momentos de mayor oferta de las categorías diamétricas requeridas en cada tipo de explotación. Hoy los rangos diamétricos que más abundan van de 2.5 a 10 cm.
El impacto generado toma una mayor significancia si recordamos que desde antes de la explotación, la especie dominante era Rhizophora sp. Es decir que la extracción maderera abarcó todo el ecosistema de manglar de la ensenada impactando la especie más importante entre los productores primarios del sistema.
Los árboles más desarrollados aportan más a la cobertura total del bosque, por lo tanto la disminución en la complejidad estructural del manglar afectó considerablemente la fauna asociada debido a la consecuente disminución en el aporte de nutrientes y a la disminución de los nichos ecológicos.
Los pobladores coinciden en que ha mermado de forma importante la oferta de la fauna asociada al manglar (piangua, alacho, meón, etc.). Sin embargo para conocer la manera como se ha afectado el flujo energético y las redes tróficas por la explotación maderera del manglar, se necesita hacer un estudio más detallado que ponga en evidencia la disminución de las poblaciones de los organismos asociados al manglar y sus causas directas.
Conclusiones
La extracción comercial de madera realizada en el ecosistema de manglar de la ensenada de Virudó en los últimos 50 años ha afectado de forma marcada su complejidad estructural (Anexo 1).
Después de haber sido fuertemente estresado, el ecosistema de manglar de la ensenada de Virudó solo se presentaron cambios en zonación en suelos que tenían un proceso avanzado de sedimentación y consolidación.
Más que un cambio en zonación lo que se ha presentado en el ecosistema de manglar es una autosucesión del género Rhizophora debido a un retroceso del proceso sedimentológico en la mayor parte de la ensenada (Figura 2, 4, anexo 1).
El manglar ribereño de la ensenada de Virudó muestra una gran capacidad de regeneración. El mangle rojo ha sido la más afectada de las morfoespecies del género Rhizophora y esto se ha debido tanto a la presión selectiva como al retroceso del proceso sedimentológico.
La extracción comercial de madera ha tenido un impacto más significativo sobre el ecosistema de manglar que la actividad artesanal de las comunidades. La industria del mangle ha modificado los patrones culturales de los habitantes de la ensenada de Virudó trayendo consigo un desequilibrio en la interfase cultura-ecosistema.
La valoración del saber de tradicional es necesaria para el estudio del ecosistema de manglar porque además de aportar información valiosa sobre ecología permite tener una perspectiva histórica de la dinámica del ecosistema. El uso y valoración del conocimiento tradicional es importante por que valora el ser y el que hacer de los habitantes de las comunidades, y rompe con la condición histórica de ignorancia que se atribuye a las culturas tradicionales.
Recomendaciones. Dado que los pobladores coinciden en afirmar que la oferta de la fauna asociada al manglar (piangua, alacho, meón, etc.) ha mermado de manera significativa, algo que implica conocer la manera como se ha afectado el flujo energético y las redes tróficas por la explotación maderera del manglar, se recomienda realizar investigaciones que ausculten y obtengan tales evidencias, así como las causas directas asociadas a la disminución de las poblaciones de los organismos asociados al manglar.
Es fundamental implementar labores de monitoreo del ecosistema de manglar en las parcelas estudiadas para hacer seguimiento tanto a la recuperación del mismo como a los actores tensionantes que aún se mantienen. En particular se debe prestar especial atención a las zonas contiguas al natal para evaluar cómo se está desarrollando la dinámica sucesional en estas áreas.
Agradecimientos. El autor agradece a todos los compañeros del grupo de trabajo de INICIATIVA BAUDÓ por su Historia de Vida: Intensa, fraterna, esperanzadora, solidaria, comprometida, consecuente y lúdica. Julio, Nancy y Swamy, Chepa, Leo, Mono, Germán, Andreita, Osquitar, La mona y Marino; todos parte del mismo equipo. A mis Padres por su complicidad. Aurita, Manuel y Héctor Ibargüen, el “viejo Hector”, por la paciencia y todas las comodidades que me brindaron para hacer este trabajo. A Eterio Ibargüen por su amistad, su comprensión y por haberme permitido aprender de su sabiduría, sin él este trabajo difícilmente se habría hecho. Al Padre Pedro Pablo, Teresa Duque y José Hurtado por el apoyo, ayuda y respaldo local. A Robert Tulio González y Humberto Alvarez por su apoyo constante y asesoría. A los Consejos Comunitario de Virudó y Cuevita, y a todos sus hombres y mujeres: Tio Máximo, Jacobo, Gabriel Cordoba, Patterson, Herminía, Tita, Guamba, Gordito, James, Luz Marina y todos los demás poseedores de la misma esperanza. Al Proyecto “Ecogenética de Poblaciones de Manglar en el pacífico colombiano” por el apoyo a la segunda salida de campo. Un agradecimiento muy especial al Doctor Luis Carlos Pardo-Locarno, por rescatar este trabajo de los anaqueles, y por creer en su importancia para el conocimiento y conservación de los manglares del Pacífico.
 Anexo 1
Estructura del manglar de
Virudó, chocó Sector del estero Bocón
Anexo 1
Estructura del manglar de
Virudó, chocó Sector del estero Bocón
Transecto 1: en esta zona se destaca la
importancia del mangle blanco tanto en términos generales como en las etapas de
brinzal, latizal y fustal; con IVI´s de 1.71, 2.35, 1.66 y 1.39
respectivamente. En tanto la densidad total es de 203 ind/0.1 ha.
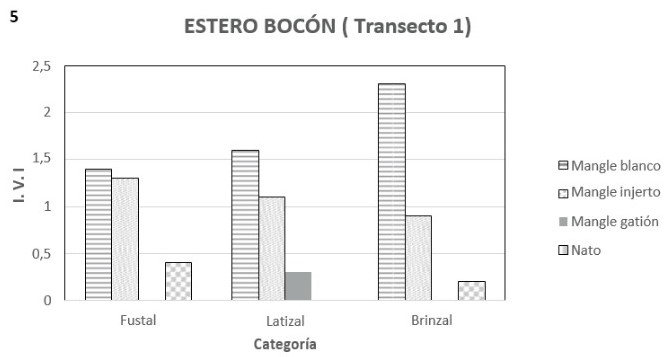
En la etapa de brinzal con el mangle blanco se encuentra asociado el mangle injerto y el nato, con IVI´s de 0.44 y 0.19, y densidades de 18 y 3 ind/0.1 ha respectivamente (figuras 4, 5 y anexo D); estas densidades muy inferiores a la de mangle blanco que tiene 96 ind/0.1 ha. Las alturas en esta categoría van de 5 a 12 metros. En la etapa latizal con el mangle blanco se encontró mangle injerto y mangle gatión con IVI´s de 1.11 y 0.22 y densidades de 22 y 4 ind/ha respectivamente (figuras 5 y 6); en tanto que la densidad total es de 79 ind./ha. Finalmente la etapa fustal mostró la presencia también de mangle injerto y nato con IVI´s de 1.22 y 0.37, y densidades de 2 y 1 ind/0.1 ha respectivamente (figuras 5 y 6). La densidad total encontrada fue de 7 ind/0.1 ha. La altura máxima alcanzada en esta etapa fue de 40 metros. Es de anotar que a pesar de la mayor densidad de mangle blanco, tanto el nato como el mangle injerto tienen una cobertura relativa mayor, con 0.64 y 0.60 respectivamente, contra 0.32 del mangle blanco.
La salinidad intersticial en marea baja fue de 16.36. En este transepto se observó gran cantidad de embriones de mangle, sobre todo concentrados en los primeros cien metros desde la orilla.
Igualmente en esta parte, en comparación con la segunda mitad del transecto, no había presencia de algas en las raíces de los mangles y la presencia de piacuil (Littorina zebra y L. scabra) era mucho menor, al igual que las bromelias, que sólo se les encontraba en los árboles más grandes.
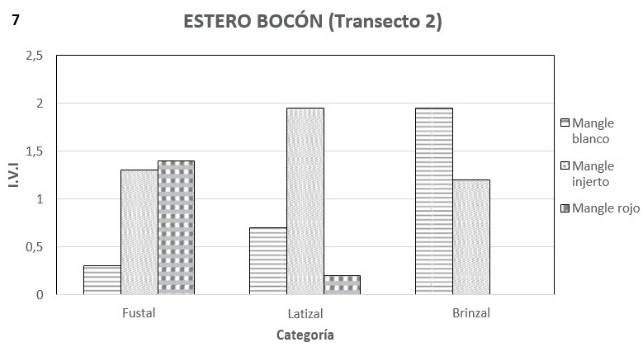
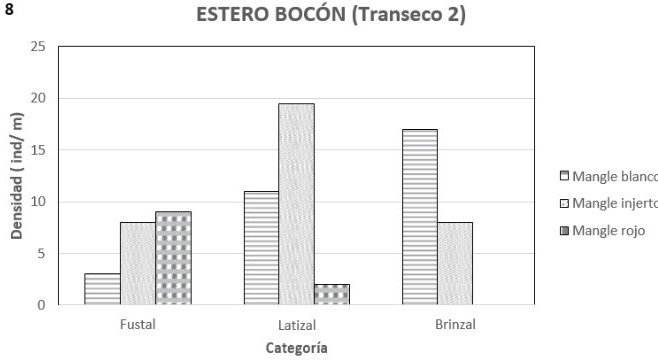
Transepto 2: a diferencia del transecto anterior, solo se encontraron asociados el mangle blanco, el injerto y el rojo y la morfoespecie más importante fue el mangle injerto con un IVI de 1.51, seguido por el blanco con 0.91 y finalmente el rojo con 0.62. La densidad total es de 78 ind/0.1 ha.
En la etapa de brinzal se destaca el mangle blanco con un IVI de 1.94 y una densidad de 17 ind/0.1ha, seguido por mangle injerto con un IVI de 1.05 y una densidad de 8 ind/0.1 ha (figuras 7 y 8). La altura mostrada por este brinzal varió entre 5 y 10 metros. La etapa latizal muestra al mangle injerto como la morfoespecie más importante con un IVI de 1.94 y una densidad de 19 ind/0.1 ha, le siguen en importancia el mangle blanco con 0.80 de IVI y con 11 ind/0.1 ha y finalmente el mangle rojo con un IVI 0.21 y 1 ind/0.1 ha (figuras 7 y 8). La altura de estos latizales osciló entre 7.5 y 14.5 metros. La categoría diamétrica de fustal mostró al mangle rojo coma la morfoespecie más destacada, seguida del injerto y del blanco con IVI´s de 1.19, 1.17 y 0.53 respectivamente (figura 7). El mangle rojo fue también la más densa con 10 ind/0.1 ha y de último el mangle blanco con 3 (figura 8). La densidad total fue de 19 ind/0.1 ha. La altura de los fustes más altos alcanzó los 40 metros.
 Figura
5
Análisis por Categoría para las especies más
importantes del transecto en el Estero Bocón.
Figura
5
Análisis por Categoría para las especies más
importantes del transecto en el Estero Bocón.
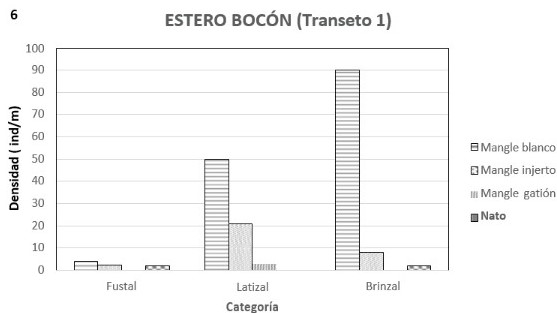 Figura 6
Análisis por categoría de la Densidad para las especies más
importantes en el transepto 1.
Figura 6
Análisis por categoría de la Densidad para las especies más
importantes en el transepto 1.
Con relación al transepto anterior se
observó que en este había mayor abundancia de fustales y menor de brinzales y
latizales. La salinidad intersticial en marea baja fue 18.08 partes por mil. El
suelo en este transecto estaba más consolidado y caminar en el manglar no se
dificultaba por la menor presencia de raíces. Igualmente a partir de la mitad del
transecto hacia adentro del manglar se observó mayor presencia de algas y de
piacuil. En esta misma parte se observó evidencia de tala de pilote,
aproximadamente 10 pilotes fueron abandonados. La marea en esta zona alcanza
una altura de 1 metro y en momentos de la quiebra no se alcanza a inundar.
 Figura
7
Análisis por Categoría para las especies más
importantes del transecto 2.
Figura
7
Análisis por Categoría para las especies más
importantes del transecto 2.
 Figura
8
Análisis por Categoría de la Densidad para las
especies más importantes en el transecto 2.
Figura
8
Análisis por Categoría de la Densidad para las
especies más importantes en el transecto 2.
Sector del Estero Peñitas
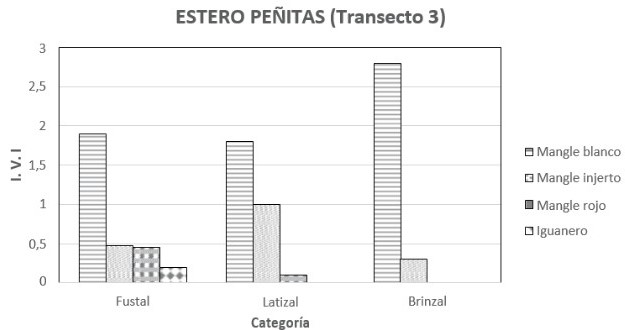
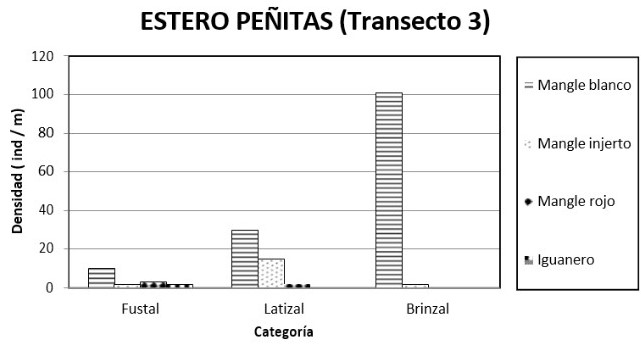
Transecto 3: en términos generales el mangle blanco fue la morfoespecie más importante con un IVI de 1.38 siendo la densidad relativa la que más aportó con 0.82. Le siguieron el mangle injerto y el mangle rojo con IVI´s de 0.91 y 0.70, y densidades de 19 y 10 ind/0.1 ha respectivamente. Estas dos últimas tuvieron coberturas relativas casi dos veces mayor que la del mangle blanco con 0.41 y 0.35 respectivamente. La densidad total fue de 166 ind/0.1 ha.
En la categoría brinzal solo se hicieron presentes dos morfoespecies de Rhizophora. El mangle blanco fue la más destaca con un IVI de 2.83 seguida por mangle injerto (figura 9). La densidad del mangle blanco fue de 102 ind/0.1 ha, en tanto que la densidad total fue de 103 ind/0.1 ha (figura 9). Las alturas en esta categoría oscilaron entre los 4 y los 7 metros. La etapa latizal mostró la asociación de las tres morfoespecies de mangle, siendo el blanco el más importante con un IVI de 1.82 y el mangle rojo el menos importante con 0.21 (figura 9). La densidad del mangle blanco fue de 31 ind/0.1 ha y la del rojo de 1 ind/0.1 ha (figura 10). La densidad total fue de 43 ind/0.1 ha. Las alturas oscilaron entre 7 y 14 metros. Finalmente en la categoría diamétrica de fustal, se destacó el mangle rojo seguido por mangle injerto y por mangle blanco. Los IVI´s fueron de 1.36, 1.27 y 0.36 respectivamente (figura 9). En cuanto a la densidad se encontró 9,8 y 2 ind/0.1 ha en el mismo orden anterior (figura 10). La altura de los fustales no superó los 37 metros de altura.
 Figura
9
Análisis por Categoría para las especies más
importantes del transepto 3.
Figura
9
Análisis por Categoría para las especies más
importantes del transepto 3.
 Figura
10
Ídem análisis categoría de la Densidad para las
especies más importantes en el transepto
Figura
10
Ídem análisis categoría de la Densidad para las
especies más importantes en el transepto
Al igual que los anteriores transeptos en la primera mitad se observó una gran densidad de plántulas de mangle (entre 60 y 100 por parcela) con DAP inferior a 2.5 cm. La salinidad intersticial en marea baja fue de 16.76 partes por mil. En esta primera mitad se dificultaba mucho caminar. Sin embargo, había presencia de algas en las raíces de los mangles, las cuales alcanzaban una altura de 60 cm, lo que muestra el nivel de inundación de la marea. A partir de la segunda mitad del transecto el bosque se hizo mucho menos denso, había menor cantidad de plántulas de mangle, igualmente el manglar se hizo más fangoso. Si bien el piacuil y el puyudo (Thais kiosquiformis) estuvieron presentes en todo el transepto, la parte interior mostró mayor presencia de estos caracoles. Como en el anterior transepto se observó evidencia de tala de pilote, varios palos fueron abandonados después de cortarlos. Los árboles más grandes fueron los únicos donde se observaban bromelias.
Transepto 4: en comparación con los transeptos anteriores, éste mostró una mayor riqueza de especies, siete en total (mangle blanco, injerto, rojo, nato, piñuelo y pelaojo). La más destacada fue la morfoespecie mangle blanco con un IVI de 2.10 y una densidad de 108 ind/0.1 ha, en tanto que la densidad total fue de 122 ind/0.1 ha. Al mangle blanco le siguieron en importancia el mangle injerto y el mangle rojo con IVI´s de 0.34 y 0.18 respectivamente, y una densidad de 2 ind/0.1 ha cada uno. La especie menos importante fue el nato con un IVI de 0.062 y con la presencia de un solo individuo en 0.1 ha.
 Figura 12
Ídem, análisis por categoría
de la Densidad para las especies más importantes en el transepto
Figura 12
Ídem, análisis por categoría
de la Densidad para las especies más importantes en el transepto
En la categoría de brinzal no se
presentaron el mangle injerto, el rojo, ni el gatión. El más importante fue el
mangle blanco con un IVI de 2.25 y una densidad de 42 ind/0.1 ha (figura 10 y
11). La densidad total fue de 49 ind/0.1 ha.
La categoría latizal mostró también al mangle blanco como la
morfoespecie más destacada con un IVI de 2.64 y una densidad de 58 ind./0.1 ha
(figuras 10 y 11), en tanto que la densidad total fue de 61 ind/0.1 ha. Ahora
la etapa fustal nuevamente mostró al mangle blanco como la más destacada con un
IVI de 1.94 (figura 10) y una densidad de 9 ind./0.1 ha (figura 11). Le
siguieron el mangle injerto y el rojo con IVI´s de 0.44 y 0.43, y densidades de 1 y 2
ind/0.1 ha respectivamente.
En comparación con el transecto anterior el
suelo en este era más inestable. En el borde del estero se observó tala de
árboles con dimensiones cercanas a las utilizadas para leña. La salinidad
intersticial promedio en marea baja fue de 19.22 partes por mil. Al igual que
otros transectos anteriores en la segunda mitad el bosque era menos denso y
también se observó evidencia de tala de pilote. En la primera mitad había
presencia de muchas plántulas de mangle.
Sector del estero El Encanto
Transepto 5: en esta zona se presentaron un total de cinco especies: Mangle
blanco, injerto, rojo, piñuelo y pelaojo. La más destacada fue la morfoespecie
mangle blanco con un IVI de 1.44 y una densidad de 129 ind/0.1 ha,
encontrándose una densidad total de 151 individuos. La segunda más importante
fue el mangle injerto con 0.77 de IVI y 2 ind/0.1 ha de densidad. Es de anotar
que la mayor cobertura relativa fue para el injerto con 0.41, la del blanco fue
de 0.31. La especie que menos aportó fue el mangle rojo con un IVI de 0.19 y
una densidad de 3 ind/0.1 ha.
 Figura
13
Análisis por Categoría para las especies más
importantes del transepto 5.
Figura
13
Análisis por Categoría para las especies más
importantes del transepto 5.
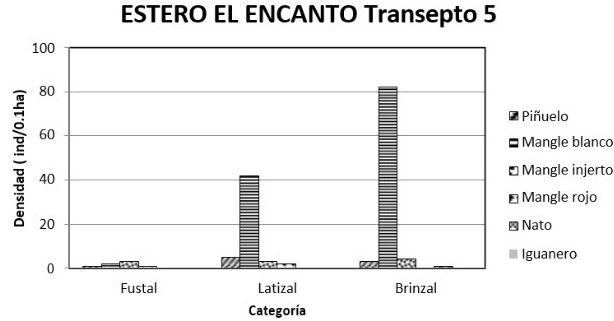 Figura 14
Ídem análisis por categoría de
la Densidad para las especies más importantes de ese transepto
Figura 14
Ídem análisis por categoría de
la Densidad para las especies más importantes de ese transepto
La categoría diamétrica brinzal mostró al mangle blanco como la morfoespecie más importante con un IVI de 2.34 y una densidad de 82 ind/0.1 ha (figuras 13 y 14) habiendo 91 ind/0.1 ha de densidad total. Después encontramos al mangle injerto como la segunda más importante con 0.32 de IVI y 2 ind/0.1 ha. Nuevamente en la etapa latizal el mangle blanco vuelve a ser la morfoespecie más importante teniendo 1.92 de IVI y 44 ind/0.1 ha de densidad (figuras 13 y 14). Le siguen el mangle injerto y el piñuelo, ambos con 0.42 de IVI y 4 y 5 ind/0.1 ha respectivamente. El total de individuos para 0.1 ha fue 57. Finalmente la categoría diamétrica fustal con solo 19 ind/0.1 ha de densidad total destacó al mangle injerto como la morfoespecie más importante con 1.45 de IVI y 10 ind/0.1 ha (figuras 13 y 14). Le siguieron el mangle blanco con 0.56 y el piñuelo con 0.43 de IVI. Cada uno de ellos con 3 ind/0.1 ha de densidad.
En términos generales este transecto mostró las mismas características cualitativas que los anteriores transeptos, es decir mayor densidad en la primera mitad y con evidencias de tala en la parte posterior. La salinidad promedio fue de 20.35 partes por mil. No se observó la presencia de bromelias en los árboles más grandes. Las algas sin embargo se hicieron presentes en las raíces de todo el transepto, alcanzando una altura de 1.20 metros en la raíz.
Transecto 6: a diferencia de todos los anteriores transectos, este destacó al piñuelo como la especie más importante con 1.07 de IVI y una densidad de 126 ind/0.1 ha. Le sigue el mangle rojo con 0.68 de IVI y 12 ind/0.1 ha, sin embargo la cobertura relativa de éste último es superior a la del piñuelo. El mangle rojo tiene 0.44 y el piñuelo 0.39. Después de estos dos en orden de importancia encontramos al nato con 0.34 de IVI y 12 ind./0.1 ha de densidad (anexo C).
La etapa de brinzal mostró nuevamente al piñuelo como la especie más importante con 2.00 de IVI y 52 ind/0.1 ha (figuras 14 y 15). La densidad total fue de 65 ind/0.1 ha. En orden decreciente de importancia aparecieron el mangle blanco con 0.39 y el injerto con 0.27 de IVI y densidades de 6 y 3 ind/0.1 ha respectivamente. Ahora, en la etapa latizal el piñuelo con 1.83 de IVI y 62 ind/0.1 ha de densidad fue la especie más importante (figuras 14 y 15). En segunda instancia el nato aparece con 0.52 de IVI y 6 ind./0.1 ha de densidad. En la categoría fustal el piñuelo sigue siendo la más importante con 1.21 de IVI (figura 14), seguido por el mangle rojo con 1.12. La densidad de estos es 12 y 7 ind/0.1 ha respectivamente (figura 15). En última instancia aparece el mangle blanco como la morfoespecie que menos aporta con 0.13 de IVI y 1 individuo en 0.1 ha.
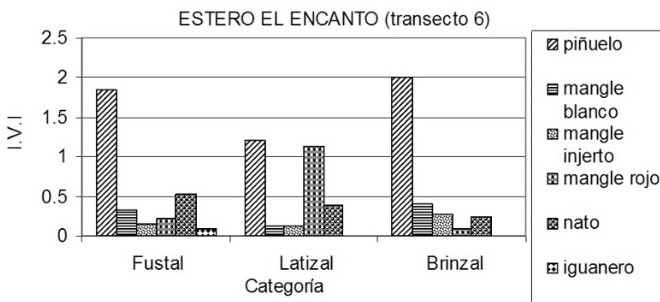 Figura
14.
Análisis por Categoría para las especies más
importantes del transecto 6.
Figura
14.
Análisis por Categoría para las especies más
importantes del transecto 6.
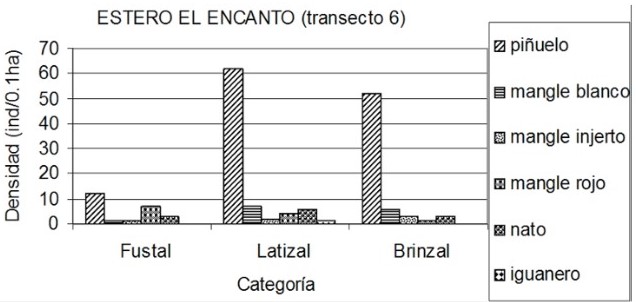 Figura 15.
Análisis por categoría de la Densidad para las especies más
importantes en el transecto 6.
Figura 15.
Análisis por categoría de la Densidad para las especies más
importantes en el transecto 6.
A diferencia de los otros transectos, el suelo en éste es firme y se inunda solo con las mareas más altas del mes. La salinidad promedio fue de 13.46 partes por mil. En las parcelas donde se encontraban los natos el suelo era más firme, diferente eran las parcelas donde abundaba más el mangle, en estas las botas se hundían. Según los valores obtenidos en este transepto se podría decir que este tipo de ecosistema de manglar es lo que la comunidad vecina llama piñuelal, que puede ser una rodal de piñuelo o un bosque asociado con otras especies pero donde predomina éste árbol.
Una característica de todos las zonas donde se hizo transeptos es la marcada densidad de la categoría brinzal y en contraste la baja densidad de la categoría fustal. En cuanto a las especies dominantes, también es claro que el mangle blanco juega el papel más destacado a excepción del último transepto donde domina el piñuelo. En segundo lugar de dominancia aparece el mangle injerto.
En las figuras anteriores vemos como en la medida en que la categoría diamétrica es más pequeña aumenta la importancia del mangle blanco, caso contrario al mangle injerto que en la medida en que la categoría evaluada es mayor encuentra una mayor importancia.
Comparando ahora entre los transectos (figuras 16 y 17), es evidente que el transepto 1 en el
Estero Bocón, tiene un marcado nivel de intervención. La cobertura de este transepto es la menor de todas con 0.77 m²/0.1 ha, valor muy distante del 3.28 m²/ 0.1 ha del transecto 6 en El Encanto. El encanto a su vez muestra la mayor cobertura, la segunda mayor densidad con 171 ind/0.1 ha
(la primera es la del transecto 1 con 203 ind/0.1 ha) y la mayor cobertura de la categoría fustal. El transepto 6 por lo tanto es la zona menos intervenida, hecho explicable por la poca utilización comercial que ha tenido el árbol de piñuelo. El transecto 1 del estero Bocón al contrario, además de mostrar la menor cobertura muestra la mayor densidad, evidencia de un estado temprano de regeneración (predomina la categoría de brinzal), congruente con la historia de explotación (estero históricamente más explotado) y por su mayor cercanía al pueblo de Virudó.
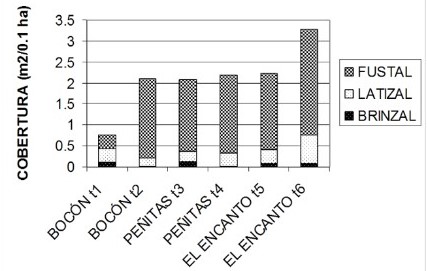 Figura 16.
Análisis comparativo de la
cobertura total entre los transectos y la proporción de las categorías
diamétricas para cada uno de ellos
Figura 16.
Análisis comparativo de la
cobertura total entre los transectos y la proporción de las categorías
diamétricas para cada uno de ellos
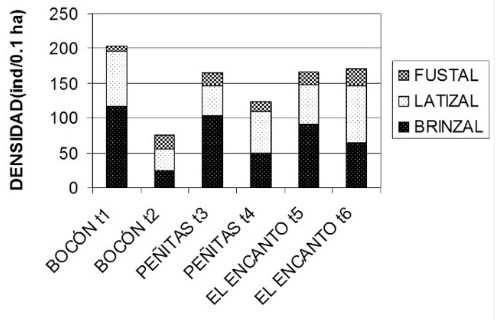 Figura 17.
Análisis comparativo de la densidad total de árboles entre
transectos y de las proporciones de las categorías diamétricas para cada uno de
ellos
Figura 17.
Análisis comparativo de la densidad total de árboles entre
transectos y de las proporciones de las categorías diamétricas para cada uno de
ellos
Sobre los perfiles de vegetación (Figuras 18, 19; 20; 21; 22 y 23) podemos decir que además de mostrar lo arriba dicho, es decir la mayor densidad de las categorías diamétricas de brinzal y latizal y la dominancia del mangle blanco, muestra que en los transectos 1,3,4 y 5 la parte más cercana al estero concentra los árboles más pequeños. Esto mismo lo observamos en la figura 24, donde vemos que la categoría brinzal se concentra en las primeras parcelas, y hacia la parte posterior de los transectos las distintas categorías tienen proporciones similares.
Ahora considerando a cada una de las categorías como una especie para averiguar como es la distribución en el espacio según el índice de Morista, encontramos que tanto la categoría de brinzal (índice de Morista=1.38, chi²=187.3, 9 g.l, X²cal0.05=16.919) como la de latizal (índice de Morista=1.19, chi²=76.48, 9 g.l, X²cal 0.05=16.919) muestran una distribución agregada. Al contrario la categoría de fustal con un índice de 1.1 muestra una distribución uniforme (figura 24).
Otro aspecto interesante que se observa en los perfiles es la mayor presencia, en las últimas parcelas de los transectos, de fustes de mangle rojo. Esto mismo lo muestra la figura 25, donde en las últimas parcelas, es decir a partir de 120 metros de la orilla del estero la probabilidad de encontrar un mangle rojo aumenta. En este gráfico no se tomó en cuenta el transecto 6, puesto que este se consideró como piñuelal, es decir un ecosistema con una matriz vegetal diferente a la del propio manglar. Aplicando el índice de morista para determinar que tipo de distribución muestra cada una de las morfoespecies de mangle (blanco, injerto y rojo), encontramos que el mangle rojo tiene un índice de 2.38, significativamente distinto a 1 (chi2=46.28, X²cal 0.05=16.919), igualmente el mangle blanco con 1.3 y significativamente diferente a 1 (chi2=74.8, X²cal 0.05=16.919). La distribución de estas dos morfoespecies es agregada, el blanco hacia la orilla del estero y el rojo hacía el fondo del manglar. Al contrario de estas dos morfoespecies el mangle injerto con un índice de 1 muestra una distribución uniforme. Lo anterior nos sugiere que evidentemente existen diferencias en la zonación de estas morfoepecies.
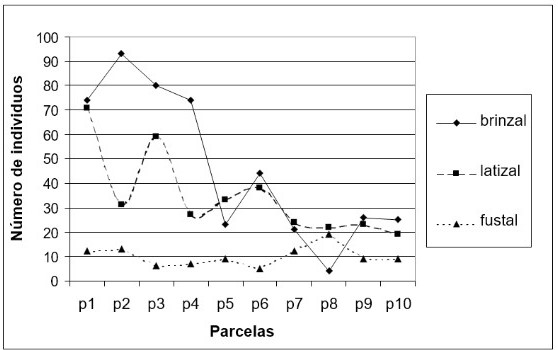 Figura 24.
Variación del número total de
individuos de las distintas categorías diamétricas en cada una de las parcelas
de todos los transeptos.
Figura 24.
Variación del número total de
individuos de las distintas categorías diamétricas en cada una de las parcelas
de todos los transeptos.
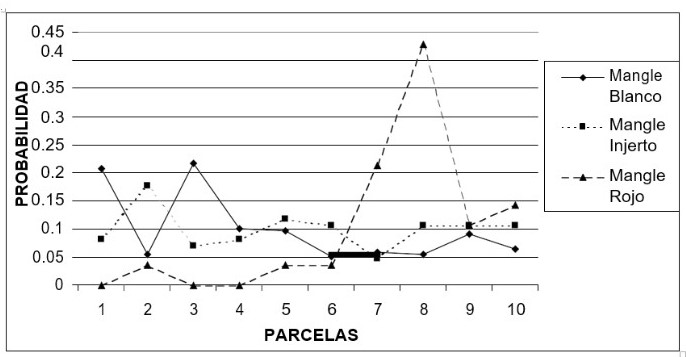 Figura 25.
Distribución de la probabilidad de encontrar cada una de las tres
morfoespecies de mangle en las parcelas de los transeptos
Figura 25.
Distribución de la probabilidad de encontrar cada una de las tres
morfoespecies de mangle en las parcelas de los transeptos
Ahora bien, con respecto a la participación en número de individuos de estas mismas tres morfoespecies en las categorías diamétricas de latizal y fustal, se observa (figura 26) que para la categoría de fustal la mayor participación corresponde a la del mangle rojo con 36.84% seguido por mangle injerto con 35.52% y mangle blanco con 27.63 %. Este orden se invierte para la categoría de latizal, donde el mangle blanco aparece con 74.45 %, después viene el injerto con 21.89 % y finalmente el rojo con tan solo 3.63 %. Este gráfico revela una aparente ausencia de regeneración del mangle rojo, mientras el mangle blanco evidencia una marcada capacidad para regenerarse. Tal situación podría explicarse por las características propias de cada una de estas morfoespecies. El mangle blanco y el injerto, al tener un crecimiento más acelerado que el de mangle rojo, se imponen sobre los embriones del mangle rojo.
Con respecto al dosel, los perfiles (figuras 18, 19, 20, 21, 22 y 23) muestran que si bien en todos los transectos hay presencia de árboles con alturas iguales o mayores a 30 metros, este estrato no es continuo; hay extensos claros en los cuáles domina un estrato inferior que no supera los 15 metros y que se presenta de forma continua. A excepción del transecto 6, donde se observa mayor heterogeneidad de estratos, los demás transectos solo presentan dos estratos, uno superior (más de 30 metros de altura) y uno inferior (entre 5 y 15 metros).
 Figura 26.
Porcentaje de individuos de Mangle Blanco, Injerto y Rojo presentes
en las categorías de fustal y latizal
Figura 26.
Porcentaje de individuos de Mangle Blanco, Injerto y Rojo presentes
en las categorías de fustal y latizal
Teniendo en cuenta la figura 27 donde se
muestra la abundancia total según rango diamétrico del conjunto de
morfoespecies de Rhizophora spp.
(Mangle blanco, injerto y rojo), es evidente el vacío de los rangos diamétricos que van de 15 a 40
centímetros. Este vacío de rangos diamétricos se puede explicar a partir de la
historia de explotación del pilote, que se detuvo tan solo hasta hace 7 años. El
leve aumento del número de individuos hacia las mayores categoría se puede
deber a los arboles que por su dificultad para ser descortezados no fueron
derribados, sumados a aquellos que no fueron utilizados porque en el momento de
determinado periodo de explotación su diámetro no correspondía a los diámetros
utilizados.
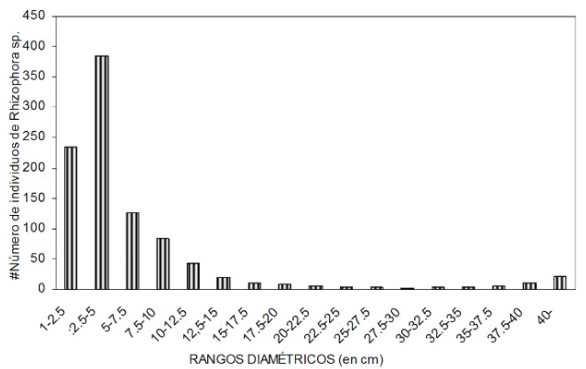 Figura 27.
Distribución del número de
individuos de Rhizophora spp. para distintas categorías diamétricas.
Figura 27.
Distribución del número de
individuos de Rhizophora spp. para distintas categorías diamétricas.
Referencias
Aceves-Lozano, J. E. (1999). La historia
oral y de vida: Del recurso técnico a la experiencia de investigación. En:
Galindo-Cáceres, J. (cord.). Investigación
en sociedad, cultura y comunicación. (pp. 207-276). México: Addison Wesley
Longuman.
Báez-Valdez, R. E. y González-Rondón, O.
(1980).Tabla de Volúmenes para Rhizophora mangle por el método de los
coeficientes mórficos empíricos. En: Vegas-Vélez, M. (ed.). Memorias Seminario
Sobre Estudio Científico e Impacto Humano en el ecosistema de Manglar. (pp.
80-88). Cali, Colombia; Montevideo, Uruguay: Universidad del Valle; UNESCO.
Referencias
Bateson, G. (1976). Pasos hacía la ecología de la Mente. Buenos Aires, Argentina:
Ediciones Carlos Lohlé
Bejarano, A. Satizabal, A. y Zapata, F. (1992). Estructura del bosque y
granulometría del suelo en un manglar de ribera de la costa Pacífica
Colombiana. En: Memorias VII Seminario
Nacional Ciencias Tecnológicas Marinas Tomo I. (pp. 120-128). Santa Marta,
Colombia.
Blanco-Libreros, J. F. y Cantera, J. R.
(1995). Patrones
estructurales de algunos manglares de la bahía de Buenaventura (Pacífico
Colombiano) Y las condiciones hidrológicas y de intervención humana que los
determinan. En: Cantera, J. Y Restrepo, J. (eds). Delta del río San Juan Bahías de Málaga y Buenaventura Pacifico
Colombiano, Tomo II. (pp.33-58).
Bohorquez, C. A. y Prada, M. C. (1984).
Efecto de los
derrames de hidrocarburos sobre el bosque de manglar en la Bahía de Cartagena a
finales de 1992. En: Yanine-Diaz D. (ed.). Memorias
Seminario Regional sobre la ordenación integrada de las zonas de manglar,
FAO/Inderena Cali (Valle) y Tumaco (Nariño) Colombia, Nov. 19-23, Tomo I.
(pp.155-165).
Referencias
Botero, L. y Mancera-Pineda, J. E. (1996). Síntesis de los cambios de
origen antrópico ocurridos en los últimos 40 años en la ciénaga de Santa
Marta (Colombia). Revista de la Academia
Colombiana de Ciencias 20 (78): (pp. 465-474).
Referencias
Bravo-Pazmiño, H. (1998). Diversidad cultural y manglares del Pacífico
Colombiano. Bogotá, Colombia: Ministerio del Medio Ambiente.
Referencias
Cantera, J. R. y Arnaud, P. (1995). Structure et distrubution des
associations d´arbres de mangrove de deux baies de la cöte pacifique de
Colombie: Málaga et Buenaventura. En: Restrepo, J. y Cantera, J. (eds.) Delta del Río San Juan Bahías de Málaga y
Buenaventura Pacífico colombiano. Tomo l. (pp. 245-286).
Day, J. W.; Hall, C. A. S.; Kempp, W. M. y Yañez-Arancibia, A. (1989). Estuarine
Ecology. New York, USA: Jahn Wiley & Sons
Referencias
De Las Salas, G. y Von Hildebrand, P. (1980). Esbozo de un plan de investigaciones para
el ecosistema de manglar Ciénaga Grande de Santa Marta Canal del Dique,
Colombia. En: VegasVélez, M. (ed.). Memorias
Seminario Sobre Estudio Científico e Impacto Humano en el ecosistema de Manglar, (pp. 378-393). Cali, Colombia; Montevideo,
Uruguay: Universidad del Valle; UNESCO.
Referencias
Delgado-Florez, A. y Sanchez-Sanchez, R.
(1965). Estudio preliminar de los manglares de la costa
Pacífica Colombiana (Cabo Corrientes- Río Baudó).
(Tesis de Grado). Universidad Distrital “Francisco José de Caldas”, Bogotá,
Colombia.
Referencias
Friedemann, N. S. (1989). Crielie criele son: Del Pacífico Negro. Bogotá Colombia: Planeta.
Referencias
Gómez, J. A. (1996). Algunas observaciones etnobotánicas
en tres comunidades Embera del Medio
Atrato Antioqueño. Cespedesia, 21(67): 197-205
Referencias
Gonzalez, J. L.; Marín L. C.; Martinez, J.
O. y Carvajal, J. H. (1998). Geomorfología y aspectos erosivos del Litoral
Pacífico Colombiano. Colombia: Publicaciones geológicas especiales del
INGEOMINAS.
Referencias
Henao-Londoño, C. I. (1996). Interpretación
de la tradición oral: Estrategia metodológica para abordar estudios
etnobiológicos. Cespedesia 21(67):
291-30.
Referencias
Hernandez-Camacho, J. I.; Von Hildebrand,
P. y Alvarez-León, R. (1980).
Problemática del manejo de manglares con especial referencia al sector
occidental de la Ciénaga Grande de Santa Marta, Magdale, Colombia. En:
Vegas-Velez, M. (ed.). Memorias Seminario
Sobre Estudio Científico e Impacto Humano en el ecosistema de Manglar. (pp.
364-383). Cali, Colombia; Montevideo, Uruguay: Universidad del Valle; UNESCO.
Holdridge, L. R. (1940). Some notes on the mangrove swamps of Puerto
Rico. The Caribbean Forester 1(4):
19-29.
Horna-Zapata, R. R. (1980). Relación Suelo y Mangle (Rhyzophora mangle, Conocarpus erectus, Laguncularia racemosa,
Avicennia nitida). En: Vegas-Vélez, M. (ed.). Memorias Seminario Sobre Estudio Científico e Impacto Humano en el
ecosistema de Manglar. (pp. 195- 200). Cali, Colombia; Montevideo, Uruguay:
Universidad del Valle; UNESCO
Referencias
Lanly, J. P. y Lindquist. A. (1985). Ordenación de
manglares en Tailandia, Malasia e Indonesia. Roma: Organización de las Naciones Unidas para
la Agricultura y la Alimentación, FAO.
Lugo, A. E. (1980).
Mangrove Ecosystems: Successional or Steady State? Biotropica, 12 (2):65-72.
Lugo, A. E.; Cintron, G. y Goenaga, C.
(1980). El ecosistema
de manglar bajo tensión. En: VegasVélez, M. (ed.). Memorias Seminario Sobre Estudio Científico e Impacto Humano en el
ecosistema de Manglar. (pp. 350 - 356). Cali, Colombia; Montevideo,
Uruguay: Universidad del Valle; UNESCO.
Referencias
Mizrachi, D., R.; Pannier, y Pannier, F. (1980). Estudio de algunas
características de las estrategias de propagación e implantación de Conocarpus
erectus L. En: Vegas-Vélez, M. (ed.). Memorias
Seminario Sobre Estudio Científico e Impacto Humano en el ecosistema de
Manglar. (pp. 257260). Cali, Colombia; Montevideo, Uruguay: Universidad del
Valle; UNESCO.
Referencias
Nickerson, N. H. y Thibodeau, F. R. ( 1985). Association
between pore water sulfide concentrations and the
distribution of mangroves. Biogeochemistry
1: 183-192.
Referencias
Odum, E. P. (1969).
The strategy of ecosystem developement. Science,
164: 262-270.
Referencias
Pannier, F. y Pannier, R. (1976).
Interpretación fisiológica de la distribución de manglar en la costa del
continente Suramericano. En: Vegas-Vélez y Rojas-Beltrán, R. (eds.). Memorias Sobre el océano Pacífico
Suramericano. (pp. 537-560). Cali, Colombia: Universidad del Valle.
Prahl, H. V. (1989). Manglares de Colombia. Bogotá, Colombia: Banco de Occidente.
Referencias
Prahl, H. V.; Cantera, J. R. y Contreras,
R. (1990). Manglares y hombres del pacífico. Bogotá,
Colombia: Editorial Presencia.
Referencias
Ricaurte, C.; Lasso, J.; Castillo, L. F. y Cantera, J. R. (1992). Principales
tensores y su efecto sobre la estructura de un manglar que bordea una población
humana en Juanchaco (Bahía de Malaga). En: CCO & COLCIENCIAS (Cord. Ed). Memorias VIII Seminario Nacional De Ciencias Y Tecnología Del Mar. (pp. 457-467). Santa Marta, Colombia: COLCIENCIAS.
Sánchez, G. (1990). Guía para el laboratorio de mecánica de suelos I. Sección de Suelos,
Departamento de Mecánica de fluidos y materiales. Facultad de Ingeniería.
Cali, Colombia: Universidad del Valle.
Sánchez-Páez, H. (1994). Los manglares de
Colombia. En: Suman, D. O. (ed.). El
ecosistema de manglar en América Latina y la cuenca del Caribe: su manejo y
conservación. (pp. 21-23).
Miami, New York, USA: Rosential School of Marine and Atmospheric Science. Univ.
de Miami (Florida) & The Thinker Foundation.
Sánchez-Páez, H. R.; Álvarez-León, O. A.; Guevara- Mancera, A.;
Zamora-Guzmán, Rodríguez-Cruz & Bravo-Pazmiño, H. E. (1997). Diagnóstico y zonificación preliminar de los
manglares del Pacífico de Colombia: Bogotá, Colombia: MMA/OIMT.
Sánchez-Páez, H. O. A.; Guevara-Mancera y
Álvarez-León, R. (eds.). (1998). Conservación
y Uso
Sostenible
de los Manglares del Pacífico Colombiano. Bogotá,
Colombia: Ministerio del Medio Ambiente.
Santamaría, G. y Andrade, H. (1997).
Cartografía Social: Sistematización de una experiencia de planificación
participativa para el desarrollo sostenible. En: Memorias del seminario Participación y Medio Ambiente. (pp. 27-35).
Cali, Colombia: Universidad del Valle.
Schutter, A. (1980). Método y proceso de la investigación participativa en la capacitación
rural. Michoacán, México: CREFAL.
Serrano, L.; Botero, A.; Cardona, P. y Mancera-Pineda, J. E. (1995). Estructura del
manglar en el delta exterior del río Magdalena -Ciénaga Grande de Santa Marta-
una zona tensionada por alteraciones del equilibrio hídrico. Anales Instituto de Investigaciones Marinas.
(24): 135-164
Shaeffer-Novelli, Y. y
Cintron, G. (1986). Guía para el estudo de areas de manguezal: Estructura, funcao e flora. Sao
Paulo, Brasil: Caribbean Ecological Research.
Suman, D. O. (1994). Situación de los manglares en América
Latina y la cuenca del Caribe. En: Suman, D. O. (ed.). El ecosistema de manglar en América Latina y la cuenca del Caribe: su
manejo y conservación. (pp.
1-10). Miami, New York, USA: Rosential School of Marine and Atmospheric
Science. Univ. de Miami (Florida) & The Thinker
Foundation.





