INTRODUCCIÓN
En Panamá los barrenadores del tallo Diatraea saccharalis (Fab.), D. tabernella Dyar, Telchin licus (Durry) y Elasmopalpus lignosellus (Zeller) han sido reportados como las plagas más importantes en caña de azúcar (Esquivel, 1980; Narvaes, 1989).
Los enemigos naturales de barrenadores en tallos de caña de azúcar pueden ser nativos o introducidos. Las especies introducidas han incluido frecuentemente parasitoides (Braconidae, Tachinidae y otros) considerando su especificidad de hospederos comparada con los depredadores (Formicidae, Salticidae y otros) que son generalistas en sus hábitos alimenticios, de allí el interés en evaluar la importancia de la depredación de artrópodos sobre barrenadores puesto que frecuentemente es subvalorada (Hall, 1988; Cherry y Nuessly, 1992; Riechert y Lockley, 1984; King y Saunders, 1984; Cherry y Robert, 2009; Cherry, 2003; Richards-Haynes, 2015).
La taxa de artrópodos presentes en campos de cañas de azúcar son principalmente depredadores tales como hormigas y arañas que comen huevos, larvas y pupas de Diatraea spp. (Negm y Hensley, 1972; Ali y Reagan, 1985; Woolwine y Reagan, 2001). En estudios conducidos en América Latina, las hormigas depredadoras fueron abundantes (Jemal y Hugh-Jones, 1993). En Estados Unidos de América estudios de depredación sobre Diatraea spp fueron conducidos por la Universidad de Florida indicando la importancia de artrópodos tales como: Solenopsis invicta Buren, Pheidole dentata Mayr, Pheidole floridana Emery (Hymenoptera: Formicidae), Orius spp. (Hemiptera: Anthocoridae), Chrysopa sp. (Neuroptera: Chrysopidae), Cicindela spp. (Coleoptera: Carabidae) y diversas especies de arañas (Capinera, 2011). En Panamá ha sido reportada la depredación de huevos y larvas del barrenador gigante Telchin licus (Lepidoptera: Castiniidae) por Ectatomma tuberculatum (Ol.), Euponera cognata (Emery), Pheidole flavens (Roger), Crematogaster sp y Solenopsis geminata (F.) (Esquivel, 1983).
La cría y liberación de insectos para control biológico del barrenador del tallo Diatraea spp. en caña de azúcar es la práctica mas eficiente en América Latina (Lenteren y Bueno, 2003 ; Fuentes et al. 2012). Los parasitoides más importantes son Cotesia flavipes Cameron (Hymenoptera: Braconidae) (Badilla, 2002; Wiedenmann et al. 2003), Trichogramma spp. (Hymenoptera: Trichogrammatidae), (Flanders y Quednau, 1960; Browning y Melton, 1987) y las moscas taquinidas (Diptera: Tachinidae) Lixophaga diatraeae Towns., Billaea(Paratheresia) claripalpis Wulp., Lydella (Metagonistylum)minense (Towns.), Palpozenillia palpalis Aldr. y Diatraeophaga striatalis Towns. (Fauconnier y Bassereau, 1970; Risco, 1996) y Telenomus sp. (Hymenoptera: Scelionidae) (Fauconnier y Bassereau, 1970).
La cuantificación de la efectividad de la depredación y el parasitismo es esencial para la busqueda de candidatos adecuados para control biológico sobre barrenadores de tallos de caña de azúcar. Los métodos corrientes para medir la eficiencia del parasitismo y depredación incluyen el uso de huevos, larvas y pupas colectadas en campo de D. tabernella. Un gran número de tallos son requeridos para encontrar los estadios de Diatraea necesarios, haciendo la evaluación de parasitismo o depredación tediosa, ineficiente y de un alto costo de ejecución.
Sin embargo, estas dificultades pueden ser superadas utilizando presas centinelas tales como G. mellonella (una alternativa a insectos hospederos principales). Estas pueden ser colocadas en los campos y monitoreando que sucede con ellas podemos determinar los más importantes depredadores y parasitoides dentro de los campos de caña de azúcar. Con esta información es posible determinar si existen parasitoides o depredadores efectivos como agentes de control biológico.
Como especie centinela potencial la polilla de la cera G. mellonella es un candidato ideal. Las polillas hembras pueden ovipositar
entre 50 a 250 huevos en su período de vida (Mishra et al. 2009). La polilla de
la cera ha sido ya utilizada como hospedero para la cría masiva de parasitoides
y depredadores en diversos programas de control biológico (Ranjbar Aghdam et
al. 2015) incluyendo las crías masivas de Trichogramma
para control de Chilo sacchariphagus (Lepidoptera:
Pyralidae) (Goebel et al. 2001), L. diatraeae(Grenier et al. 1982) y Tetrastichus
howardi (Ochoa, 2013; Piñero et al. 2016) para control de D. saccharalis.
G. mellonella fue utilizada en la producción de hongo entomopatógeno Metarhizizium anisopliae en Colombia (Obando et al. 2013) y la identificación de nematodos entomopatógenos en India (Razia et al. 2011). En Colombia, esta especie también ha sido utilizada para la producción de los nematodos tales como Steinernema sp. y Heterorhabditis sp. para su potencial utilización en el manejo integrado de D. saccharalis en caña de azúcar (López-Llano y Soto-Giraldo, 2016).
El objtetivo de este estudio fue la evaluación potencial de huevos, larvas y pupas de G. mellonella como presa centinela para la evaluación de enemigos naturales de barrenadores del tallo en caña de azúcar en Panamá.
MATERIALES
Y MÉTODOS
Sitio del estudio
Este estudio fue realizado en Grupo CALESA (Compañía Azucarera La Estrella S.A.) en Natá, Panamá (N 08°17.425´; W
080°31.180´). Las observaciones fueron realizadas desde marzo 2016 a febrero
2017 (12 meses) sobre la variedad B74-125.
Cría de G.
mellonella
Para las evaluaciones se utilizaron huevos, larvas y pupas de G. mellonella. Estas producciones
fueron establecidas en el Laboratorio de Control Biológico de CALESA. Los
insectos fueron criados utilizando dieta artificial con los siguientes
ingredientes: Salvado de trigo (280 g), harina (120 g), reemplazo de leche de
ternero (160 g), miel de abeja (250 ml) (Diferentes estadios de G. mellonella son mostrados en las
figuras 1, 23).
Localización de puntos centinelas de
G. mellonella (Huevos, larvas y
pupas) en campo.
El estudio fue realizado en un campo de 9 ha dividido en
9 parcelas (cada sitio = 1 ha) (Figura
4 a). Se instaló una trampa en el centro de cada parcela (50 metros desde la
calle principal adyacente a la parcela y 30 surcos de caña desde el borde de la
parcela. La trampa consistió de un tubo (vial) Coring Eppendorf 50 conteniendo cuatro larvas y cuatro pupas (con perforaciones en la tapa) colocadas
en el medio de los tallos de caña de azúcar (Figura 4 b). Los tallos fueron
escogidos al azar y la altura del
vial dependio del tamaño del tallo. La G.mellonella
centinelas fueron revizadas una vez a la semana para verificar la presencia
de parasitoides y/o depredadores. Pupas y larvas de G. mellonella fueron llevados al laboratorio para su revisión. Al
mismo tiempo, un nuevo dispositivo centinela fue colocado con pupas y larvas
(Figura 4 b y 4 c). Revizando la depredación de huevos, 40 G.mellonella huevos fueron colocados en una etiqueta de papel con
goma (unidad) de 2 cm x 2 cm sobre una hoja de la parte superior del tallo (El ultimo internudo cerca del meristema epical). Estos fueron removidos después de siete días y regresados a el laboratorio para
revizar la mortalidad.
 Figura
1.
Larvas de G. mellonella (25 mm).
Figura
1.
Larvas de G. mellonella (25 mm).
 Figura
2.
Pupas de G. mellonella (20 mm).
Figura
2.
Pupas de G. mellonella (20 mm).
 Figura
3.
Huevos de G. mellonella (0.45 x
0.35 mm).
Figura
3.
Huevos de G. mellonella (0.45 x
0.35 mm).
 Figura 4.
Disposición en campo (Google, 2020) e
instalación de larvas, pupas y huevos sobre tallos de caña de azúcar (a, b y c).
Figura 4.
Disposición en campo (Google, 2020) e
instalación de larvas, pupas y huevos sobre tallos de caña de azúcar (a, b y c).
Todos los especimenes obtenidos de los dispositivos con larvas, pupas y huevos de G.mellonella fueron identificados y preservados en etanol al 90%.
Tasa de
depredación y parasitismo
Las tasas de depredación y parasitismo se estimaron según las siguientes fórmulas:
Tasa de depredación: Número de huevos depredados / Total huevos x 100; Número de larvas depredadas / Total larvas x 100; y Número de pupas depredadas /Total pupas x 100.
Tasa de parasitismo: Número de huevos parasitados/Total huevos x 100; (Número de larvas parasitadas/Total larvas) X 100; (Número de pupas parasitadas /Total pupas) X 100.
Porcentaje de Entrenudos barrenados
(% E.B.) y tasa de parasitismo.
Cien tallos fueron muestreados por mes (cuatro réplicas de 25 tallos molibles cada una). Estos fueron tomados de cuatro puntos dentro de 2 ha de campo. Cada tallo fue medido longitudinalmente para obtener la altura (cm) y abiertos longitudinalmente para contar el número de entrenudos barrenados (E.B.) y calcular:
% E.B.: Número de entrenudos barrenados / Número de entrenudos total * 100
Tasa de parasitismo: Número de larvas parasitadas / Total larvas x 100 y/o Número de pupas parasitadas / Total de pupas x 100.
Análisis
de datos.
La
tabulación semanal de datos, el cálculo de porcentajes y los gráficos fueron
realizados con el programa Microsoft Excel 2016.
RESULTADOS
Capturas
en puntos centinelas de G. mellonella en
campo.
Se capturaron 2233 de artrópodos pertenecientes a 2 clases, 4 órdenes, 5 familias y 10 especies (Cuadro 1; Figuras 5, 6, 7 y 8).
Cuadro 1.
Artrópodos
capturados con G. mellonella en caña
de azúcar en Panamá.
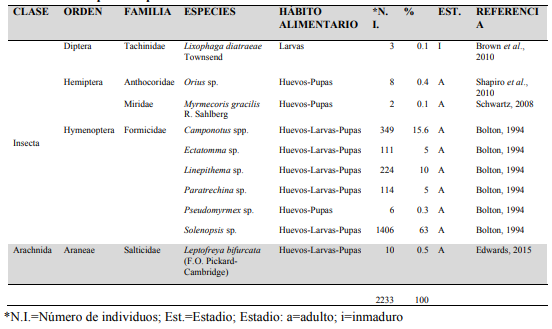 * N.I.=Número de individuos; Est.=Estadio;
Estadio: a=adulto; i=inmaduro
* N.I.=Número de individuos; Est.=Estadio;
Estadio: a=adulto; i=inmaduro
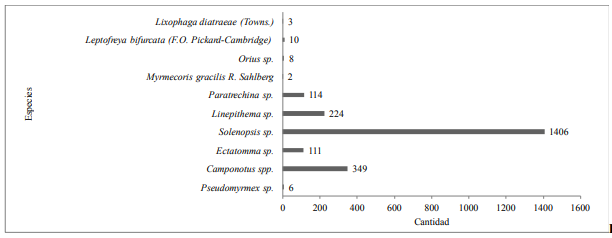 Figura 5.
Número de capturas por especie con G.
mellonella en caña de azúcar en Panamá.
Figura 5.
Número de capturas por especie con G.
mellonella en caña de azúcar en Panamá.
 Figura
6.
Camponotus spp. (10 mm).
Figura
6.
Camponotus spp. (10 mm).
 Figura
7.
Solenopsis sp. (6 mm).
Figura
7.
Solenopsis sp. (6 mm).
 Figura
8.
Pupa de G. mellonella depredada
por Camponotus spp. (10 mm).
Figura
8.
Pupa de G. mellonella depredada
por Camponotus spp. (10 mm).
 Figura
9.
Myrmecoris gracilis R. Sahlberg.
(10 mm).
Figura
9.
Myrmecoris gracilis R. Sahlberg.
(10 mm).
Tasa de depredación y
parasitismo
Para cada estadio de G. mellonella, la mortalidad debida a enemigos naturales fue detallada por huevos (95.8%), larvas (70.6%) y pupas (83.2%), con una mortalidad media de 92.6% (Cuadro 2).
Con respecto a la tasa de depredación en los cuadros 2 y 3, los resultados indican que la depredacion de huevos por Myrmecoris gracilis R.F. Sahlberg (Hemiptera: Miridae) (Figura 9) fue la mas alta, seguida por Ectatomma sp. y Camponotus spp. La depredación de larvas de G. mellonella fue principalmente debida a Paratrechina sp., seguida por Linepithema sp y Ectatomma sp. Considerando la tasa de depredación de pupas de G. mellonella, esta fue dominada por 3 especies: Myrmecoris gracilis, Linepithema sp. y Pseudomyrmex sp., dependiendo del tiempo en que las especies fueron observadas. La depredación de todos los estadios de G. mellonella fue principalmente debida a Linepithema sp., Camponotus spp. y Ectatomma sp.
Cuadro 2.
Mortalidad de G. mellonella como presa centinela en
caña de azúcar en Panamá.

Cuadro 3.
Tasa de depredación y parasitismo en
diferentes estadios de G. mellonella en
caña de azúcar en Panamá.
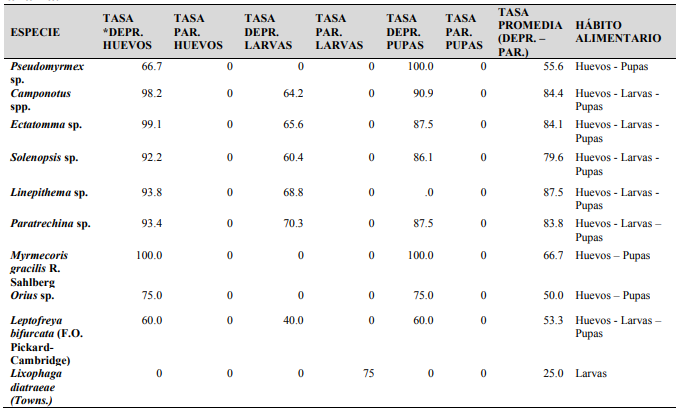 * Depr.:
Depredación; Par.: Parasitismo
* Depr.:
Depredación; Par.: Parasitismo
Porcentaje de entrenudos barrenados (% E.B.) y tasa de
parasitimo
Durante la evaluación de un periodo de 12 meses, C. flavipes (33.3%) fue encontrada parasitando larvas de D. tabernella en tallos con una altura de 221.8 cm (11 meses de edad) (Para el sitio y condiciones de este estudio). El porcentaje mas alto de entrenudos barrenados ocurrio cuando la altura de los tallos de 17.3 (4.1%), 25.9 (5%) y 45.9 cm (6.1%) (3 a 5 meses de edad) (Cuadro 4 y Figura 10).
Hubo un período activo de depredación que va principalmente desde 3 a 7 meses, correspondientes al período de crecimiento de la caña de azúcar (Cuadro 4 y Figura 10).
Cuadro 4.
Presencia de
hormigas y el parasitoide C. flavipes durante
el período de crecimiento de caña de azúcar en Panamá.
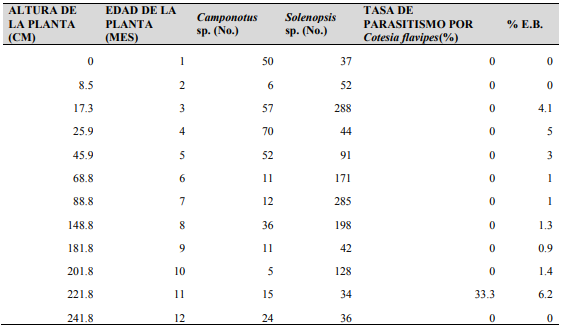
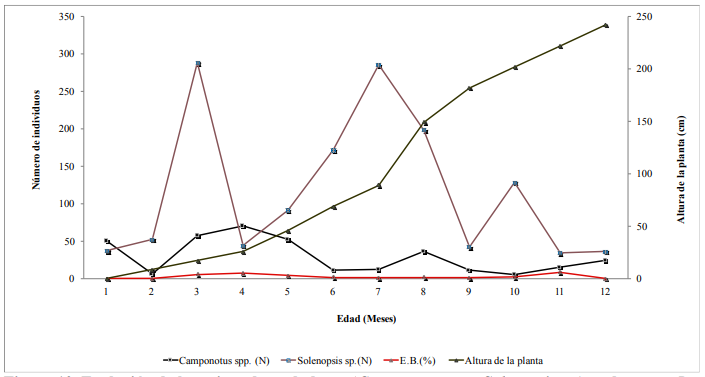 Figura 10.
Evolución de hormigas
depredadoras (Camponotus spp.-Solenopsis sp.) vs daño por D. tabernella (% E.B.) en relación a la
altura del tallo (cm) en Panamá.
Figura 10.
Evolución de hormigas
depredadoras (Camponotus spp.-Solenopsis sp.) vs daño por D. tabernella (% E.B.) en relación a la
altura del tallo (cm) en Panamá.
DISCUSIÓN
Capturas en puntos centinelas de G. mellonella en campo.
Formicidae fue la familia predominante comprendiendo 6
especies, dentro de las cuales las especies más capturadas fueron Solenopsis sp. (63%) y Camponotus spp. (15.6%). El impacto de
las hormigas como depredadores sobre barrenadores del tallo de la caña de
azúcar como el caso por ejemplo de Diatraea spp. ya había sido demostrado por ciertas investigaciones realizadas en diferentes regiones de América (Negm y
Hensley, 1972; Ali y Reagan, 1985; Jemal y Hugh-Jones, 1993; Woolwine y Reagan,
2001), lo que indica que el uso de puntos centinelas de G. mellonella en campo tiene una utilidad práctica para establecer
las poblaciones, especies y potencial impacto de dichos depredadores.
Tasa de depredación y parasitismo
La eficiencia de depredación de especies de hormigas sobre huevos, larvas y pupas de G. mellonella coincidieron con otros registros de depredación establecidos sobre barrenadores del tallo como Diatraea spp. (Negm y Hensley, 1972; Ali y Reagan, 1985; Woolwine y Reagan, 2001).
Huevos y pupas de G. mellonella no atrajeron parasitoides durante el período de observación, mientras el estadio de larva fue parasitado (75%) por la mosca taquinida L. diatraeae (Diptera: Tachinidae), parasitoide relacionado a Diatraea spp. en estudios realizados (Fauconnier y Bassereau, 1970; Risco, 1996).
Se destaca que se encontró el depredador M. gracilis, que es un chinche que mimetiza con hormigas del género Formica y fue reportada depredando afidos, pequeños insectos y huevos de insectos (Wachmann et al. 2004). Las especies de hormigas (Formicidae) Ectatomma sp., Camponotus spp., Linepithema sp., Pseudomyrmex sp., fueron reportadas como depredadores D. saccharalis y otros artrópodos sobre plantaciones de caña de azúcar (Oliveira et al. 2012; Bailly-Maitre et al. 2012). Paratrechina fulva fue reportada como agente de control para hormigas cortadoras de hojas (Atta sp.); sin embargo, esta especie puede desplazar entomofauna nativa particularmente otras hormigas por consiguiente decreciendo la biodiversidad del ecosistema (Arcila et al. 2002).
Porcentaje de entrenudos barrenados
(% E.B.) y tasa de parasitimo
El parasitoide C. flavipes que fue encontrado dentro de los entrenudos de los tallos, es según los estudios realizados el más importante parasitoide de Diatraea spp. (Badilla, 2002; Wiedenmann et al. 2003).
El más alto número de Solenopsis sp. fue capturado con G. mellonella cuando la altura de los tallos fue de 17.3 cm (3 meses de edad), que coincide cuando el nivel de daño de D. tabernella fue 4.1% E.B. y también cuando el más alto número de Camponotus sp. capturados coincide cuando la altura del tallo fue 25.2 cm (4 meses de edad) cuando el nivel de daño de D. tabernella fue de 5 % E.B. La tasa de parasitismo más alta de C. flavipes sobre larvas de D. tabernella coincidió con el pico de 6.1% E.B. por D. tabernella cuando la altura fue de 221.8 cm (11 meses).
La importancia de la depredación de artrópodos sobre barrenadores que había sido frecuentemente subvalorada, ha retomado importancia durante los últimos años sobre todo por los aspectos relacionados ha establecer un equilibrio agroecológico dentro de los campos de cultivos (Cherry y Nuessly, 1992; Riechert y Lockley, 1984; Cherry y Robert, 2009; Cherry, 2003; Richards- Haynes, 2015).
CONCLUSIONES
Los enemigos naturales de barrenadores de tallos
reportados fueron principalmente de la familia Formicidae con 6 especies. Las
especies dominantes fueron: Solenopsis sp.
(63%) y Camponotus spp. (15.6%). La presencia de hormigas como depredadores en campos de caña de azúcar
evidencia que existe una red trófica presente y necesita ser preservada. Existe
un uso potencial de G. mellonella como
centinelas para estudios de entomofauna principalmente buscando determinar la
presencia e impacto de depredadores.
Agradecimientos
SENACYT y Grupo CALESA (Compañía Azucarera la Estrella, S.A.) por proveer el soporte técnico y
logístico. Guillermo Ramirez (Departamento de Campo), Angela María Fuentes
Gómez (Laboratorio de Biotecnología) y Amed Ramos (Técnico de Campo) por su colaboración. Expresamos igualmente nuestra gratitud hacia William White
(USDA ARS, Louisiana) por revisar el manuscrito.
REFERENCIAS
Ali,
A.D. y Reagan, T.E. (1985). Spider inhabitants of sugarcane ecosystems in
Louisiana an update.
Proceedings
of the National Academy of Sciences, 48,
18–22.
Arcila, A.M., Gómez, L.A. y Ulloa-Chacón, P. (2002). Immature Development and Colony Growth of Crazy ant Paratrechina fulva Under Laboratory Conditions. Sociobiology, 39(2), 1-15.
Badilla, F. (2002). Un programa exitoso de control biológico de insectos plaga de la caña de azúcar en Costa Rica. Manejo Integrado de Plagas y Agroecología (Costa Rica), 64, 77-87.
Bailly-Maitre, J., Goebel, R. y Vercambre, B. (2012). Evidence of the role of predatory ants in natural pest control in banana-sugarcane rotation systems. Entomologie faunistique – Faunistic Entomology, 65, 49-68
Bolton, B. (1994). Identification Guide to the Ant Genera of the World. Cambridge, Massachusetts: Harvard University Press.
Brown, B.V.,
Borkent, A., Cumming, J.M., Wood, D.M., Woodley, N.E. y Zumbado, M. (2010).
Manual of Central
American Diptera Vol. 2. NRC Research Press.
Browning, H.W. y Melton, C.W. (1987). Indigenous and exotic trichogrammatids (Hymenoptera: Trichogrammatidae) evaluated for biological control of Eoreuma loftini and Diatraea saccharalis (Lepidoptera: Pyralidae) borers on sugarcane. Environmental entomology, 16(2), 360-364.
Capinera, J.L. (2011). Sugarcane Borer, Diatraea saccharalis (Fabricius) (Insecta: Lepidoptera: Pyralidae). Retrieved from http://edis.ifas.ufl.edu/in374.
Cherry, R.H. y Nuessly, G.S. (1992). Distribution and Abundance of Imported Fire Ants (Hymenoptera: Formicidae) in Florida Sugarcane Fields. Environmental Entomology, 21(4), 767– 70.
Cherry, R. (2003). The Effect of Harvesting and Replanting on Arthropod Ground Predators in Florida Sugarcane. Florida Entomologist, 86(1), 49–52.
Cherry, R. y Robert, G. (2009). The Effect of Harvesting and Replanting on Arthropod Ground Predators in Florida Sugarcane. University of Florida. IFAS Extension (pp 1–3).
Edwards, G.B. (2015). Freyinae, a major new subfamily of Neotropical jumping spiders (Araneae: Salticidae). Zootaxa, 4036, 1-87.
Esquivel, R. (1980). Basic studies on sugarcane resistant varieties to the giant borer (Castnia licus Drury) in Panama. Entomology Newsletter, International Society of Sugarcane Technologists, 8, 8-9.
Esquivel, R.E.A. (1983). Effective control of the giant mothborer Castnia licus Drury in Panama, utilising biological-cultural methods. Entomology Newsletter, International Society of Sugar Cane Technologists, 14, 6-7.
Fauconnier, R. y
Bassereau, D. (1970). Les Enemis Animaux de la Canne à Sucre. La Canne à Sucre.
Techniques
Agricoles et Productions Tropicales. Collection dirigée par René Coste.
Flanders, S.E. y Quednau, W. (1960). Taxonomy of the genus Trichogramma (Hymenoptera, Chalcidoidea,
Trichogrammatidae). Entomophaga, 5(4), 285-294.
Fuentes, F., Ferrer, F. y Salas, J. (2012). Reseña Histórica del Control Biológico En Centroamérica y El Caribe. Revisión de logros del Control Biológico en la Región neo-tropical con énfasis a los principales países del Caribe. Editorial Académica Española, 200 pp.
Goebel, R., Tabone, E., Rochat, J. y Fernandez, E. (2001). Biological Control of the Sugarcane Stem Borer Chilo sacchariphagus (Lep: Pyralidae) in Réunion Island: Current and Future Studies on the Use of Trichogramma spp. Proceedings of South African Sugar Technologists Association, 75, 171-174.
Grenier, S., Barthelemy, S. y Bonnot, G. (1982). Dynamique de la maturation des œufs chez le parasitoıde Lixophaga diatraeae (Diptera, Tachinidae) élevé dans un hôte de substitution Galleria mellonella (Lepidoptera, Pyralidae). Reproduction Nutrition Développement, 22(3), 523-535.
Hall, D.G. (1988). Insects and Mites
Associated with Sugarcane in Florida. Florida
Entomologist,
71(2), 138–150.
Jemal, A. y Hugh-Jones, M. (1993). A review of the red
imported fire ant (Solenopsis invicta Buren)
and its impacts on plant, animal, and human health. Preventive Veterinary Medicine, 17(1–2), 19- 32.
King, A.B.S. y Saunders, J.L. (1984). Las Plagas Invertebradas
De Cultivos Anuales Alimenticios En America Central. Overseas Develpment Administration, ODA/CATIE/TDRI. 182 pp.
Lenteren, J.C. y Bueno, V.H.P. (2003). Augmentative
biological control of arthropods in Latin America. BioControl, 48(2),
123-139.
López-Llano, R. A. y Soto-Giraldo, A. (2016). Aislamiento
de Nematodos Entomopatógenos Nativos en Cultivos de Caña Panelera y Pruebas de
Patogenicidad Sobre Diatraea saccharalis (Lepidoptera:
Crambidae). Boletín Científico. Centro de Museos. Museo de Historia Natural, 20(2),
114-123.
Narvaes, L. (1989). Caña de Azúcar. En: K. Andrews y J. Quezada (Eds.), Manejo Integrado de Plagas
Insectiles en la Agricultura: Estado Actual y Futuro. Honduras: Escuela
Agrícola Panamericana. El Zamorano.
Negm, A.A. y Hensley, S.D. (1972). Role of predaceous
arthropods of the sugarcane borer Diatraea
saccharalis (F.) in Lousiana. Proceedings of the International Society of
Sugar Cane Technologists Fourteenth Congress, New Orleans, Louisiana, October
22 November 5, 445-453.
Obando, J.A., Bustillo, A.E., Castro, U. y Mesa, N.C. (2013).
Selección de cepas de Metarhizium
anisopliae para el control de Aeneolamia
varia (Hemiptera: Cercopidae). Revista
Colombiana de Entomología, 39(1),
26-33.
Ochoa, A. (2013). Algunos aspectos etológicos y efectividad
de Tetrastichus howardi (Olliff)
(Hymenoptera: Eulophidae), en el control de Diatraea
saccharalis (Fab.) (Lepidóptera : Pyralidae) en áreas forrajeras de la
Empresa Azucarera Majibacoa.Tesis de Master en Pastos y Forrajes. Cuba:
Universidad De Matanzas Camilo Cienfuegos Estación Experimental De Pastos y
Forrajes Indio Hatuey.
Oliveira, R.D.F.,
Almeida, L.C., Souza, D.R., Munhae, C.B., Bueno, O.C. y Morini, M.S.D. C.
(2012).
Ant
diversity (Hymenoptera: Formicidae) and predation by ants on the different
stages of the sugarcane borer life cycle Diatraea
saccharalis (Lepidoptera: Crambidae). European
Journal of Entomology, 109(3),
381-387.
Piñeyro, N.G., Fagundes Pereira, F., Gomes Borges, F.L.,
Rossoni, C., Souza Silva, A. y Oliveira Kassab, S. (2016). ¿Multiplicar Tetrastichus howardi (Hymenoptera:
Eulophidae) en la oruga de la seda afecta su biología?. Acta Biológica Colombiana, 21(1),
189-193.
Ranjbar Aghdam, H., Yousefi Porshokouh, A. y Sedighi, L.
(2015). Temperature-dependent life table parameters of Galleria mellonella (L.) (Lepidoptera: Pyralidae). Journal of Crop Protection, 28(4), 727-738.
Razia, M., Padmanaban, R., Karthik, Raja, R., Chellapandi, P. y Sivaramakrishnan. (2011). Monitoring
entomopathogenic nematodes as ecological indicators in the cultivated lands of
Karur District, Tamil Nadu: a survey report. Electronic Journal of Biology, 7(1), 16-19.
Richards-Haynes, C. (2015). Indigenous Natural Enemies of the Sugarcane Stem Borer (Diatraea spp) their Relationship and Importance in the Guyana Sugarcane
Eco-System. Guyana Sugar Corporation Inc, Agricultural Research Department No.
11. Recuperado de: http://www.jamaicasugar.org/WIST2004/PDF%20files/Papers/Paper14.PDF
Riechert, S. y Lockley, T. (1984). Spiders as Biological
Control Agents. Annual Review of
Entomology, 29, 299–320.
Risco, S.H. (1996).
Éxitos históricos de taquinidos y braconidos en el control biológico de Diatraea
en cana de azúcar cultivada en América. Revista Peruana de Entomología, 39,
85-90.
Schwartz, M.D. (2008). Revision of the Stenodemini with a
Review of the Included Genera (Hemiptera: Heteroptera: Miridae: Mirinae). Proceedings of the Entomological Society of
Washington, 110(4), 1111-1201
Shapiro, J.P., Shirk, P.D., Kelley, K., Lewis, T.M. y
Horton, D.R. (2010). Identity of two sympatric species of Orius (Hemiptera: Heteroptera: Anthocoridae). Journal of Insect Science, 10, 189.
Wachmann, E., Melber, A. y Deckert, J. (2004).
Cimicomorpha: Microphysidae (Flechtenwanzen), Miridae (Weichwanzen).
Neubearbeitung der Wanzen Deutschlands, Österreichs und der deutschsprachigen
Schweiz. Die Tierwelt Deutschlands. Wanzen. Band 2 [in German]. Goecke &
Evers, Keltern. 288 pp.
Wiedenmann, R. N., Smith, J. W. y Rodríguez-del-Bosque, L. A.
(2003). Host suitability of the New World stalkborer Diatraeaconsiderata for three Old World Cotesia parasitoids. BioControl,
48(6), 659-669.
Woolwine, A.E. y
Reagan, T.E. (2001). Potential of Winter Cover Crops to Increase Abundance of
Solenopsis invicta (Hymenoptera:
Formicidae) and Other Arthropods in Sugarcane.
Environmental
Entomology, 30(6), 1017-1020.
Notas de autor
1 Universidad
de Montpellier, Escuela Doctoral GAIA, Francia. randy.atencio@gmail.com
2 CIRAD
(Centro de Cooperación Internacional en la Investigación Agronómica para el
Desarrollo), unidad AÏDA (Agroecología y Manejo Sostenible de Cultivos Anuales), en Montpellier, Francia. regis.goebel@cirad.fr
3 Laboratorio de
Biotecnología, Compañía Azucarera La Estrella S.A., Panamá (Grupo CALESA). abby.guerra@grupocalesa.com
3 Laboratorio de
Biotecnología, Compañía Azucarera La Estrella S.A., Panamá (Grupo CALESA). silvia.lopez@grupocalesa.com




