1. Introduction
Early weaning of calves at two months of age significantly improves the reproductive performance of primiparous and multiparous cows in poor body condition at the beginning of the service period(1)(2)(3), but it challenges the nutrition of the early-weaned calf. During its first eight weeks of life, the calf depends greatly on its mother's milk(4), which gradually declines as lactation progresses and, with it, the specific contribution of milk in the calf's diet(5)(6). Supplementation with high energy-protein ration (me 12 mj/kg, cp 19%) on temperate pastures in British breed calves weaned at 60 days of age has been enough to achieve weight gains in the range of 0.6 to 0.7 kg/d(7)(8), and a bodyweight of around 150 kg at 6 months of age, similar to those reported for lactating calves on native pastures(7)(8). Calves' early weaning associated with lot feeding with concentrated diets allows, in addition to improving cows' reproductive rate, to significantly increase calves' live weight at 180 days of age, compared to that achieved in grazing(9), positively impacting productivity (total kilograms of weaned weight/cow exposed to bull) and economic result of the cow-calf system(10).
Given the early development stage of the early-weaned calves, protein requirements for the expression of growth potential are maximum compared to more advanced development stages(11), which leads to increasing the cp concentration in diets offered to this category. However, although it is essential to ensure the daily intake of this nutrient, excessive cp in rations is undesired, both because of the negative effect on its cost, and the environmental impact of more N excretion in feces and urine(12)(13). Although grazing animals need concentrates with 18-19% cp to complement the lower quality forage base, probably, when feeding high concentrate total mixed rations the daily supply of metabolizable protein (mp) needed to meet calves’ requirements to gain around 1.0-1.2 kg/d is achieved with less cp in the ration(14).
There is little information quantifying the response of early-weaned calves of beef breeds to the cp level in the diet, varying in the magnitude and type of response in weight gain (wg) and feed conversion efficiency, depending on animal factors, such as weaning age and breed, evaluated cp range(15)(16)(17) and concentrate level in the diet(18). On the other hand, the ability of nutritional models to predict the expected daily intake for this animal category, a fundamental input in the prediction of the net protein supply for growth, is not sufficiently validated. Pascoal and others(18) report an underestimation of the necessary cp intake by nrc (1984), compared to the observed values, given the same wg in early-weaned calves. A more precise adjustment between protein requirements and supply would contribute to improving the efficiency of the whole process.
This study aimed to evaluate the effect of increasing levels of cp in the diet on wg and feed efficiency of early-weaned Hereford calves, lot-fed with a high concentrate total mixed ration.
The hypothesis suggests that there is a level of cp contribution in the ration that optimizes the wg and conversion ratio, minimizing the N excretion.
2.
Material and methods
The
experiment was carried out in Cassinoni Experimental
Station (eemac)
of the Agronomy College (Paysandú, 32° 38´ S; 58° 04´W) in the summer-autumn of
2013. Twenty-four open-air pens were used for individual housing (10 m2),
with 30% of shaded area and provided with a drinker with ad libitum
water supply and a feeder each.
2.1
Animals, experimental design and treatments
Twenty-four
Hereford calves castrated at birth and early-weaned at 80±10 days of age
(90.1±13.6 kg) were randomly assigned to one of four feeding treatments, after
a 14-day transition period to solid diet. Four totally mixed rations were
formulated (tmr,
82% concentrate, 18% roughage) differing in cp
concentration: 12, 15, 18, or 21% (dry matter basis). The increase in cp content was generated by
substituting ground corn grain for soybean meal, keeping the urea and the other
ingredients constant, resulting in iso-energetic rations (me=12.0 ± 0.03 mj/kg; Table1).
Table 1
Ingredient and nutritional composition of the
experimental rations
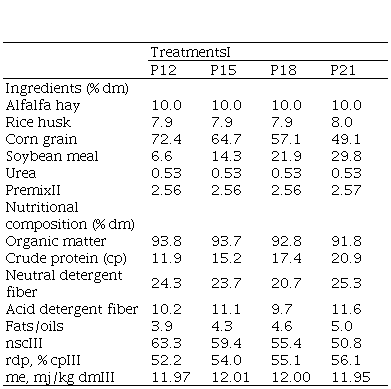 I. Crude protein level in the ration:
12, 15, 18 and 21%, dry matter (dm)
basis. II. Premix including: CaCO3 (57.2%), NaCl (17.1%), Rumensin-10% monensin (2.3%); Zoodry feedlot
(vitamin-mineral supplement) (11.7%); Beef-sacc
yeasts (11.7%). III. Values based on bcnrm(14), rdp: rumen degradable protein; nsc: non-structural
carbohydrates; me: metabolizable
energy
I. Crude protein level in the ration:
12, 15, 18 and 21%, dry matter (dm)
basis. II. Premix including: CaCO3 (57.2%), NaCl (17.1%), Rumensin-10% monensin (2.3%); Zoodry feedlot
(vitamin-mineral supplement) (11.7%); Beef-sacc
yeasts (11.7%). III. Values based on bcnrm(14), rdp: rumen degradable protein; nsc: non-structural
carbohydrates; me: metabolizable
energy
|
TreatmentsI
|
|
P12
|
P15
|
P18
|
P21
|
|
Ingredients
(% dm)
| | | | |
|
Alfalfa
hay
|
10.0
|
10.0
|
10.0
|
10.0
|
|
Rice husk
|
7.9
|
7.9
|
7.9
|
8.0
|
|
Corn grain
|
72.4
|
64.7
|
57.1
|
49.1
|
|
Soybean
meal
|
6.6
|
14.3
|
21.9
|
29.8
|
|
Urea
|
0.53
|
0.53
|
0.53
|
0.53
|
|
PremixII
|
2.56
|
2.56
|
2.56
|
2.57
|
|
Nutritional
composition (% dm)
| | | | |
|
Organic
matter
|
93.8
|
93.7
|
92.8
|
91.8
|
|
Crude
protein (cp)
|
11.9
|
15.2
|
17.4
|
20.9
|
|
Neutral
detergent fiber
|
24.3
|
23.7
|
20.7
|
25.3
|
|
Acid
detergent fiber
|
10.2
|
11.1
|
9.7
|
11.6
|
|
Fats/oils
|
3.9
|
4.3
|
4.6
|
5.0
|
| nscIII |
63.3
|
59.4
|
55.4
|
50.8
|
| rdp, % cpIII |
52.2
|
54.0
|
55.1
|
56.1
|
| me, mj/kg
dmIII |
11.97
|
12.01
|
12.00
|
11.95
|
Calves were group fed during the transition to solid diet according to the protocol reported by Simeone and Beretta(7). After this process, the calves, already assigned to their treatment, were transferred to the experimental facilities, and for three weeks they gradually adapted to the individual lot housing and were introduced to their experimental diets. By the end of this period animals reached ad libitum intake and the initial lw was recorded (107.2 ± 21.5 kg).
The calves were treated against internal and external parasites with injectable 1% ivermectin (Ivomec® from Laboratorio merial), and constantly monitoring their health status from then on.
During the 12-weeks experimental period, food was offered ad libitum in two daily meals of the same amount (8 am and 7 pm), adjusting the supply through bunk reading, so that residual food was not less than 10% of the offered.
2.2
Measurements and sampling
Live weight (lw) was recorded every 14 days without fasting, before the first meal of the day. The dry matter intake (dmi) was estimated daily as the difference in dry weight between the offered feed and the residual food present in the feeder, which was weighed and discarded. Samples of the offered feed were taken weekly to determine the dm content, while individual leftovers were sampled every 15 days. Samples were dried in a forced-air stove at 60 °C to constant weight, then ground to 2 mm and preserved for further chemical analysis.
Rump height(19), Longissimus dorsi muscle area (rea), and subcutaneous dorsal fat thickness (dft) were measured at the beginning and end of the experimental period. These last two were determined by ultrasonography at 12.a and 13.a rib height, placing the transducer laterally in that area, and specific software for beef breed evaluation (BioSoft Tool Box II, Biotronic) processed the images.
The tmrs apparent digestibility was estimated in vivo in week 11 of the experiment, in two animals per treatment, through the total collection of feces(20). They were collected for three consecutive days, directly from the floor of each lot, by directly observing the animals during the day and collecting the night production in the following morning before the first feeding. Fresh feces were weighed every 24 hours and one subsample per animal was dried for dm content determination, and then preserved for further chemical analysis. Blood and urine samples were taken on day 4 of week 11 from the same animals 4 hours after the first feeding of the day. The urine samples were taken by spontaneous urination according to the technique described by Santos and others(21), keeping a subsample of 10 ml per animal diluted in 0.036 N sulfuric acid, so the pH is lower than 3. Blood samples were extracted by a puncture on the jugular vein and put in a test tube containing edta as anticoagulant, immediately centrifuged at 5000 rpm for 15 minutes, and the plasma conserved at -15 °C. In week 11, daily samples were taken from the supplied and leftover food. Samples were dried in a forced-air stove at 60 °C to constant weight to determine dm content, and preserved for further chemical analysis.
2.3 Chemical analysis
The
chemical composition of the offered and leftover feed was analyzed on composite
samples by treatment for the entire experimental period, determining ash
content(22), neutral detergent fiber using α-amylase and correcting by ash contamination (aNDFom), acid detergent fiber (adf)(23) and cp (total N*6.25)(22). Ash, cp and aNDFom
concentration were also analyzed on composite samples of food and feces per
animal corresponding to week 11 of the experimental period, and urea concentration
was determined in plasma (bun)
and urine (uun)
samples(22).
2.4 Calculated variables
The feed conversion ratio was calculated as the ratio between the daily average dmi (kg/d) and the wg (kg/d). The apparent digestibility of dm (dmd, %), organic matter (omd), cp and ndf were calculated as the difference between daily intake of each fraction (fi) and the product of feces concentration (fc) by daily feces production (fp): D=(fi g/d – fc% × fp g/d) / fi g/d. The fc was estimated from nutrient concentration of the (nc) in the offered feed (of) and in the leftovers (cf, g/d = of kg/d × nc g/kg - leftovers kg/d × nc g/kg).
The urinary excretion of N (nu, g/d) was estimated based on the prediction equation reported by Kohn and others(24), from bun (g/L) concentration and the urea elimination rate through urine (cr, L filtered blood per day/kg lw): nu g/d.kg lw = cr × bun. The considered cr value was 1.3 ± 0.12 L/d.kg lw reported for cattle by the same authors.
To interpret the observed response, the contribution of metabolizable protein (mp) and metabolizable energy (me) in the different treatments was estimated from the bcnrm model (Beef Cattle Nutrient Requirements Model)(14). The observed values of dmi and lw were used as inputs, and the tabular values of degradability of cp and the carbohydrates provided by the model were considered for the ingredients of the diet (under the assumption that they present the same amino acid composition and fermentation pattern as the ingredients used in tmrs).
2.5
Statistical analysis
The
experiment was analyzed using linear models corresponding to a completely
randomized design considering the animal as the experimental unit. Variables
with repeated measures over time (lw and dmi) were analyzed using the Mixed Procedure of sas (sas Institute
Inc., Cary, nc),
including the initial lw
as covariate. The wg
was analyzed according to a linear mixed model of slopes heterogeneity of lw as a
function of time, and the slopes of the fitted lines were compared by simple
contrasts. Variables without repeated measures over time were analyzed using
the sas glm
procedure. When the treatment effect was significant, the linear and quadratic
effects associated with the level of inclusion of cp in the tmr were evaluated. An effect was considered
significant when the probability of type I error was lower than 5%, and it was considered a trend when it was lower than 10%.
3.
Results
3.1
Animal growth
The
effect of treatments on the variables that describe growth and development of
calves is presented in Table 2.
Table 2
Effect of protein level in diet on growth and
development parameters of early-weaned calves (84 days post-weaning)
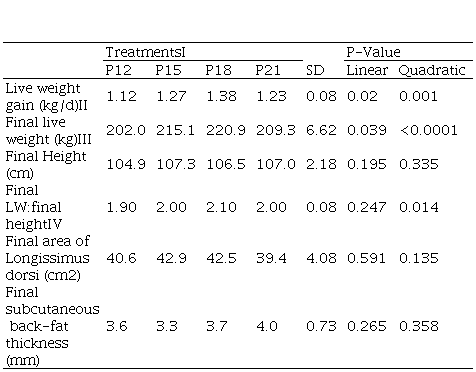 II. Increasing levels of crude protein in diet 12, 15, 18 and 21% of ration in dry matter basis. II. WG, kg/d= -0.0083x2 + 0.2897x - 1.167; R² = 0.93. III. LW, kg= -0.6861x2 + 23,565x + 17,515; R² = 0.97. IV. LW:Height = -0.0056x2 + 0.1967x + 0.33; R² = 0.90
II. Increasing levels of crude protein in diet 12, 15, 18 and 21% of ration in dry matter basis. II. WG, kg/d= -0.0083x2 + 0.2897x - 1.167; R² = 0.93. III. LW, kg= -0.6861x2 + 23,565x + 17,515; R² = 0.97. IV. LW:Height = -0.0056x2 + 0.1967x + 0.33; R² = 0.90
|
TreatmentsI
| | |
P-Value
|
|
P12
|
P15
|
P18
|
P21
|
SD
|
Linear
|
Quadratic
|
|
Live weight gain (kg/d)II
|
1.12
|
1.27
|
1.38
|
1.23
|
0.08
|
0.02
|
0.001
|
|
Final live weight (kg)III
|
202.0
|
215.1
|
220.9
|
209.3
|
6.62
|
0.039
|
<0.0001
|
|
Final Height (cm)
|
104.9
|
107.3
|
106.5
|
107.0
|
2.18
|
0.195
|
0.335
|
|
Final LW:final
heightIV
|
1.90
|
2.00
|
2.10
|
2.00
|
0.08
|
0.247
|
0.014
|
|
Final area of Longissimus dorsi (cm2)
|
40.6
|
42.9
|
42.5
|
39.4
|
4.08
|
0.591
|
0.135
|
|
Final subcutaneous back-fat thickness (mm)
|
3.6
|
3.3
|
3.7
|
4.0
|
0.73
|
0.265
|
0.358
|
The evolution of lw showed a linear adjustment (P<0.01) and the
slope varied according to the treatment (P=0.039). The wg and final lw increased quadratically;
registering maximum values of wg and final lw (predicted from adjusted
regressions) of 1.36 kg/d and 219.4 kg for cp
levels in the tmr
of 17.5% and 18.0%, respectively (Figure 1). No differences in final rump
height were detected (P>0,10), so the lw:height ratio had the same
quadratic response as the final lw. There was no significant effect for the cp level on the rea, nor on the dft (P>0.10).
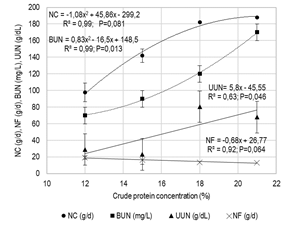 Figure 1
Nitrogen consumption (nc), total N in feces (nf), urea N
in blood (bun) and urine (uun), in
early-weaned calves fed with iso-energetic rations (me 12.0 MJ/kg) differing in crude protein concentration
Figure 1
Nitrogen consumption (nc), total N in feces (nf), urea N
in blood (bun) and urine (uun), in
early-weaned calves fed with iso-energetic rations (me 12.0 MJ/kg) differing in crude protein concentration
3.2
Intake, apparent digestibility and conversion ratio
As
the ration cp increased, dmi and N
consumption increased quadratically (P<0.01), estimating a maximum dmi of 5.75
kg/d for a cp level of 17.6%
(Table 3), while the maximum N consumption of 190 g/day was reached at cp = 21.3% (Figure 2). Intake expressed
every 100 kg of lw
did not vary with the treatment (3.84 ± 0.16 kg/100 kg lw; P>0.10), it only varied
with the experimental week (P<0.01) regardless of the type of ration
(P>0.10).
Table 3
Effect of the level of protein in the diet on consumption, apparent
digestibility and feed conversion efficiency
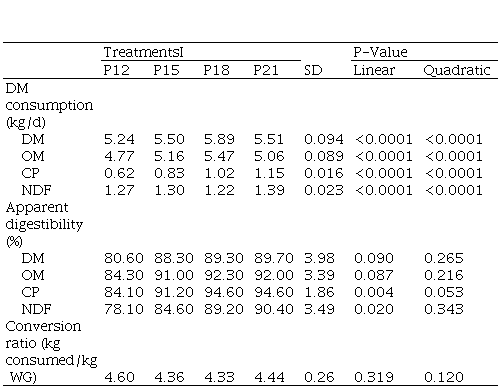 I. Crude protein level in diet 12, 15, 18 and 21% of ration in dry basis. DM Dry matter, OM: organic matter, CP: Crude protein, NDF: neutral detergent fiber, LWG: live weight gain
I. Crude protein level in diet 12, 15, 18 and 21% of ration in dry basis. DM Dry matter, OM: organic matter, CP: Crude protein, NDF: neutral detergent fiber, LWG: live weight gain
|
TreatmentsI
| | |
P-Value
|
|
P12
|
P15
|
P18
|
P21
|
SD
|
Linear
|
Quadratic
|
|
DM consumption (kg/d)
| | | | | | | |
|
DM
|
5.24
|
5.50
|
5.89
|
5.51
|
0.094
|
<0.0001
|
<0.0001
|
|
OM
|
4.77
|
5.16
|
5.47
|
5.06
|
0.089
|
<0.0001
|
<0.0001
|
|
CP
|
0.62
|
0.83
|
1.02
|
1.15
|
0.016
|
<0.0001
|
<0.0001
|
|
NDF
|
1.27
|
1.30
|
1.22
|
1.39
|
0.023
|
<0.0001
|
<0.0001
|
|
Apparent digestibility (%)
| | | | | | | |
|
DM
|
80.60
|
88.30
|
89.30
|
89.70
|
3.98
|
0.090
|
0.265
|
|
OM
|
84.30
|
91.00
|
92.30
|
92.00
|
3.39
|
0.087
|
0.216
|
|
CP
|
84.10
|
91.20
|
94.60
|
94.60
|
1.86
|
0.004
|
0.053
|
|
NDF
|
78.10
|
84.60
|
89.20
|
90.40
|
3.49
|
0.020
|
0.343
|
|
Conversion ratio (kg consumed/kg WG)
|
4.60
|
4.36
|
4.33
|
4.44
|
0.26
|
0.319
|
0.120
|
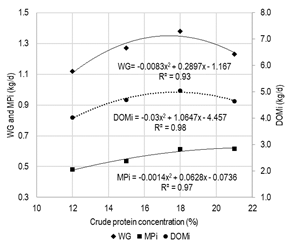 Figure 2
Weight gain (wg), metabolizable protein
intake (mpi)
and digestible organic matter intake (domi) in early-weaned calves fed
with iso-energetic rations (me
12.0 mj/kg),
differing in crude protein concentration
Figure 2
Weight gain (wg), metabolizable protein
intake (mpi)
and digestible organic matter intake (domi) in early-weaned calves fed
with iso-energetic rations (me
12.0 mj/kg),
differing in crude protein concentration
The dmd and omd tended to increase linearly with the cp content of the diet (P<0.10), as did the digestibility of ndf and cp (P<0.05). As a consequence of the intake response and the observed digestibility coefficients, digestible cp and ndf intakes increased quadratically as the cp content in the diet increased (P<0.01; Table 3), as did the digestible om intake (P<0.01; Figure 1).
The estimated daily contribution of mp based on bcnrm(14) was 481, 537, 612 and 617 g/d for cp levels in the tmr of 12, 15, 18 and 21%, respectively (Figure 1). As the proportion of soybean meal in the ration increased and the corn grain decreased, the daily contribution of rumen degradable protein in P21 doubled compared to P12 (0.33 vs 0.60 kg/d). This did not affect the synthesis of microbial protein between treatments (0.33 vs 0.38 kg/d) but modified the N balance in rumen (-9.1, 4.5, 20.5 and 34.9 g N/d, for cp 12, 15, 18 and 21%, respectively).
The feed conversion ratio, was not significantly affected by the protein level in the ration. Nonetheless, a numerical tendency to improve this indicator was observed as the protein concentration increased in the diet up to cp = 18.0%.
3.3
N in blood, feces, and urine
During
the measurement period of bun
concentration and N excretion, the daily N consumption tended to increase
quadratically with the cp level
in the diet (P <0.08), as it did during the entire experimental period.
Daily N fecal excretion decreased linearly (P<0.06), while urinary excretion
increased linearly (NU g/d= 2.29x -16.19 R² = 0.98; P<0.01). In parallel, bun and uun concentration increased
quadratically (P=0.013) and linearly (P=0.046), respectively, with the cp level in the diet (Figure 2).
4.
Discussion
4.1
Intake, digestibility, contribution of metabolizable protein and efficiency of
N use
The dmi increased quadratically when the cp level increased in the diet, maximizing at 5.75 kg/d for a level of cp=17.5% in the diet. Expressed every 100 kg of lw, the dmi was of 3.84 ± 0.84% (P>0.10), a value within the reported for this category, with ad libitum intake of concentrated rations such as the used in the present study(9). Several authors also report quadratic-type increases in dmi in response to the cp level in the diet. Veira and others(16) registered a significant increase in the dmi up to 12% of cp, stabilizing from this value. These authors evaluated the effect of the cp level in similar types of diet and calves’ age at weaning (8 to 12 weeks of age, 80 kg lw), although using a range of values lower than those of the present study (10% to 16% cp). Neville and others(17) also worked with concentrated diets (14% forage) and increasing cp levels in the diet of Hereford and Angus calves, but younger at weaning than those of the present study (48 days, varying between 21 to 60 days) and 74 kg of lw. They report a stable dmi for cp values in the diet between 14.5% and 23.5%, falling significantly when the cp increased to 28.5% (2.93 kg/d, 3.02 kg/d, 2.77 kg/d, respectively); however, N consumption increased linearly. Acayezu and others(25) also report a quadratic type response in the dmi against increasing cp levels in the diet (15% to 22%) when working with even lighter Holando calves (40 kg) but reached the maximum at higher values of cp (19%), falling significantly at 22%. On the contrary, other authors that measured the effect of the cp level in less concentrated rations (roughage>40%) found no response in dmi, both in early-weaned calves(18)(26)(27) and in finishing steers(12)(28)(29).
The regulation of voluntary dmi depends on a group of factors with short and long-term effects, being the quality of the diet one of the main involved in short-term(11) regulations, mainly in feedlot conditions. In the present study, an increase in dmi was associated with an improvement in the apparent digestibility of om, cp and ndf, a similar response to that observed by Obeid and others(28), Veira and others(16), and Neville and others(17). Likewise, this increase in dmi associated with the cp level could contribute to modifying the site of digestion for part of the diet’s starch and protein, through a positive effect on the passage rate(30).
As a result of the response in dmi and the increase in the cp concentration in the diet, the N consumption (g/kg lw0.75) increased 80% when going from 12% to 21% of cp in the diet, significantly improving the apparent digestibility of dm, om, cp and ndf. Other authors also report an improvement in the digestibility of cp(16)(25)(27), and ndf(16) by increasing the concentration of N in the diet, while other studies do not report an effect on the fiber(25)(27).
Increasing substitution levels of corn grain for soybean meal resulted in different protein concentration among the tmrs, therefore, differences in the fermentation characteristics between ingredients could have also contributed to generate differences in the synthesis of microbial biomass and the contribution of non-degradable protein in the rumen, differentially affecting the mp available in each treatment. The daily contribution of mp (kg/d), estimated based on bcnrm(14) from the observed intake and ingredient composition of each diet, increased 28% when the cp in the diet went from 12% to 21%. In parallel, the N balance calculated at ruminal level showed a linear increase, going from negative values in P12, where the contribution of rumen degradable protein was the limiting factor for the synthesis of microbial protein, to maximum levels in P21, where the balance of N in rumen represented a 57% excess according to the requirements. Furthermore, the contribution of the microbial protein to mp fell from 51.5% in P12 to 39.7% in P21, probably modifying the amino acid profile of mp as well.
From the point of view of the N use efficiency, the excess of N in the rumen resulting from the positive balance observed would increase the concentration of ruminal ammonium, its absorption through the rumen walls, and later, at a metabolic level, it would be converted into urea and excreted through urine(11). This would explain the increase observed in the bun and uun concentration in the present study, as N consumption increased (Figure 2). High N consumption has been related to high bun concentration(31) and high urinary excretion(32). Cole and others(31) report a limit bun concentration of 9 mg/dL, above which it is considered that the consumption of N would be excessive. In the present study, minimum levels of bun were registered in P12, followed by a sustained increase in bun concentration for each variation percentage unit in the cp concentration of the diet (7, 9, 12 and 17 mg/dL for ration cp levels of 12%, 15%, 18% and 21%, respectively), showing that cp levels above 15% would already be excessive (Figure 2). Kohn and others(24) report a close linear relationship between bun concentration (mg/dL) and excretion rate of N in urine (nu, g/d.kg lw) in all species. Daily nu excretion estimated for the present study was minimal in P12 (13.9 g/d), increasing 2.5 times in P21 (35.0 g/d). This elimination process is not only undesirable from an environmental point of view(12), but it would also result in an increase in energy costs for maintenance(33).
4.2 Growth and feed conversion ratio
Calves’ wg increased quadratically, registering decreasing increments up to a cp diet inclusion level of 17.5%, and a decrease from this value, accompanying the variation observed digestible om intake and mp contribution (Figure 1). Other authors also observed such response with early-weaned calves fed with concentrated diets; the optimal level differing according to age (19-20% in younger calves)(17)(25), and breed (around 14-16% in Holando calves)(15)(16).
Weight gain is determined by the contribution of energy substrates and intestinal amino acid absorption. Furthermore, protein retention will depend on the mp usage efficiency, also conditioned by the availability of energy substrates and essential amino acids that might be limiting(34). The nutritional evaluation of the diets based on the bcnrm(14) model showed that, in all treatments, the daily mp intake limited the expression of higher wg rates, which could have been achieved according to the energy intake. It is important to highlight the good fit between wg predicted by the model and the observed for the cp levels in the diet of 12%, 15% and 18% (R² = 0.97). However, this adjustment occurred when the observed intake was used, which was in all treatments, well above the one predicted by the model. The positive wg response to increases in the contribution of mp up to P18 would be a consequence of the high protein requirements during the early phase of the animal's life(11) when it registers an important growth in lean body mass, and a lower increase rate of adipose tissue, which deposits later(35). Bartlett and others(36), and Donnelly and others(37) document an increase in muscle deposition in calves as a response to the increase in the cp content of the diet, simultaneously registering an increase in water and protein content, and a decrease in fat content(37). The response observed in rea and dft, as indicators of the gain composition, was numerically consistent with the reported by the aforementioned authors. Greater muscle deposition (higher rea values) was recorded in P15 and P18, while dft was not affected; both variables presented a high negative correlation (r= -0.79). Gleghorn and others(29) reported that even when increasing cp levels in the ration did not affect the dmi of finishing steers, a quadratic increase in carcass weight was observed with a maximum of 13% cp, with no effect of the cp level on dft. wg fall as of P18 (-11%) would partly respond to the dmi drop (-6%), but it could also be due to lower efficiency in the use of mp(37), added to a change in the partition of the me consumed. The latter would be attributable to the probable increase in maintenance requirements resulting from a higher energy cost for the detoxification of N(33) excess in P21, and the consequent reduction in the proportion of consumed me available for wg.
The conversion ratio did not show statistically significant variations when varying cp concentration in the diet, which was consistent with the same type of quadratic response observed for the dmi and the wg. However, at a numerical level, more favorable values were observed as the protein in the ration increased to around 17-18%. In this range, although dmi increases, wg would do so more than proportionally. This could be associated with the positive impact of cp level on the digestibility of om, cp and ndf, the increase in mp contribution and the characteristics of amino acids absorbed regarding the requirements for wg(14). Above 18% cp, not only an improvement in cp digestibility is no longer observed (Table 3), but the metabolic cost associated with the excretion of excess N(33) would be much higher than that registered for lower cp levels in the diet, while bun concentration increased more than proportionally when the concentration of cp in the diet was of 21%.
5. Conclusions
Early-weaned
Hereford calves (80 ± 10 days-old) fed with concentrated rations (18% roughage,
me: 12.0 MJ/kg) during
post-weaning, maximized their growth when the cp
content of the diet ranged between 17% and 18%, achieving wg of 1.36 kg/d and weights
of 219 kg when leaving the feedlot. However, a lower cp level of around 15% would not significantly affect the
feed conversion and would maintain a good wg (1.31 kg/d), while
simultaneously reducing N excretion, presenting a benefit from an environmental
point of view.
References
1. Simeone A, Beretta V. Early weaning: an efficient and effective technological alternative for calf rearing in grassland cattle systems in Latin America. Cuban J Agri Sci. 2016;50(1):39-49.
2. Wettemann RP. Nutritional influences on reproduction of beef cows. In: Quintans G, Scarsi A, editors. Seminario de Actualización Técnica: Cría vacuna. Montevideo: Hemisferio Sur; 2013. p. 247-64.
3. Lusby KS, Wettemann RP, Turman EJ. Effects of early weaning calves from first-calf heifers on calf and heifer performance. J Anim Sci. 1981;53(5):1193-7.
4. Rovira J. Manejo nutritivo de los rodeos de cría en pastoreo. Montevideo: Hemisferio Sur; 1996. 288p.
5. López M, Martino G, Torres L. Evaluación del efecto del destete temporario sobre el crecimiento y comportamiento del ternero al pie de la vaca [grade’s thesis]. Montevideo (UY): Universidad de la República, Facultad de Agronomía; 2018. 96p.
6. Olaizola J, Piegas JM, Silveira X. Evaluación del uso del creep feeding sobre el comportamiento y desarrollo de los terneros [grade’s thesis]. Montevideo (UY): Universidad de la República, Facultad de Agronomía; 2016. 120p.
7. Simeone A, Beretta V. Destete precoz en ganado de carne. Montevideo: Hemisferio Sur; 2002. 118p.
8. Simeone A, Beretta V, De León M, Silvera E, Torres S, Widmaier G. Efecto del nivel de suplementación en pasturas sobre la ganancia de peso de terneros destetados precozmente. Rev Arg Prod Anim. 1997;17(Supl. 1):60.
9. Beretta V, Simeone A, Elizalde JC, Caorsi CJ, Lamarca M. Destete precoz a corral: ¿una nueva herramienta para una nueva cría? In: Una nueva cría, un nuevo engorde, una nueva ganadería. 14 Jornada anual de la Unidad de Producción Intensiva de Carne; 2012; Paysandú, Uruguay. Paysandú (UY): UPIC; 2012. p.14-27
10. Simeone A, Beretta V. Alternativas tecnológicas para la nueva cría y su impacto en el resultado físico y económico de los sistemas de producción criadores. In: Una nueva cría, un nuevo engorde, una nueva ganadería. 14 Jornada anual de la Unidad de Producción Intensiva de Carne; 2012; Paysandú, Uruguay. Paysandú (UY): UPIC; 2012. p. 38-51.
11. National Research Council. Nutrient requirements of beef cattle. 7th ed. Washington: National Academic Press; 1996. 248p.
12. Menezes A, Valadares Filho SC, Costa e Silva LF, Pacheco MVC, Pereira JMV, Rotta PP, Zanetti D, Detmann E, Silva FAS, Godoi LA, Rennó LN. Does a reduction in dietary crude protein content affect performance, nutrient requirements, nitrogen losses, and methane emissions in finishing Nellore bulls? Agric Ecosyst Environ. 2016;223:239-49.
13. Church CD. El Rumiante: fisiología digestiva y nutrición. Zaragoza: Acribia; 1988. 630p.
14. National Academies of Sciences, Engineering, and Medicine. Nutrient Requirements of Beef Cattle. 8th rev ed. Washington: The National Academies Press; 2016. 494p.
15. Hill TM, Bateman II HG, Aldrich JM, Scholotterbeck RL. Crude protein for diets fed to weaned dairy calves. Prof Anim Sci. 2008;24(6):596-603.
16. Veira DM, Macleod GK, Burton JH, Stone JB. Nutrition of the weaned Holstein calf II: effect of dietary protein level on nitrogen balance, digestibility and feed intake. J Anim Sci. 1980;50(5):945-51.
17. Neville WE, Hellwig RE, Ritter RJ, McCormick WC. Effect of diet protein level on weight gains of early weaned beef calves. J Anim Sci. 1977;44(4):687-93.
18. Pascoal LL, Eifert E, Restle J. Nível de proteína bruta para bezerros de corte desmamados a os 66 dias de idade. Rev Bras Zootec. 2000;29(5):1537-44.
19. Guidelines for uniform beef improvement programs [Internet]. 9th ed. Prairie (MS): Beef Improvement Federation; 2016 [cited 2020 Set 23]. 185p. Available from: https://bit.ly/3hTCall.
20. Lascano CE, Borel R, Quiroz R, Zorrilla J, Chávez C, Wernli C. Recomendaciones sobre metodología para la medición de consumo y digestibilidad in vivo. In: Ruiz M, Ruiz A, editors. Nutrición de rumiantes. San José (CR): IICA; 1990. p. 159-67.
21. Santos SA, De Souza JM, Valadares Filho SC, Soares A, Motta S, Ferreira AM. Balanço de nitrogênio em fêmeas leiteiras em confinamento alimentadas com concentrado à base de farelo de soja ou farelo de algodão. Rev Bras Zootec. 2010;39(5):1135-40.
22. Latimer GW, editor. Official methods of analysis. 19th ed. Gaithersburg (MD): AOAC; 2012. 2v.
23. Goering HK, Van Soest PJ. Forage fibre analyses: apparatus, reagents, procedures and some applications. Washington: USDA; 1970. 20p.
24. Kohn RA, Dinneen MM, Russek-Cohen E. Using blood urea nitrogen to predict nitrogen excretion and efficiency of nitrogen utilization in cattle, sheep, goats, horses, pigs, and rats. J Anim Sci. 2005;83(4):879-89.
25. Akayezu JM, Linn JO, Otterby DE, Hansen WP. Evaluation of calf starters containing different amounts of crude protein for growth of Holstein calves. J Dairy Sci. 1994;77(7):1882-9.
26. Cavalcante MA, Pereira O, Valadares Filho SC, Ribeiro K. Níveis de proteína bruta em dietas para bovinos de corte: consumo, digestibilidade total e desempenho produtivo. Rev Bras Zootec. 2005;34(3):711-9.
27. Tieri MP, La Manna A, Fernández E, Mieres J, Schroer F, Pérez E, Baldi F, Banchero G. Efecto de diferentes niveles de proteína y sustitución de proteína verdadera por nitrógeno no proteico (urea) en la performance y desarrollo de terneros cruza Hereford x Angus y su impacto posterior en la recría. In: Producción de carne desde una invernada de precisión [Internet]. Montevideo: INIA; 2010 [cited 2020 Set 23]. p. 23-48. Available from: https://bit.ly/3iRwdqx.
28. Obeid JA, Pereira OG, Pereira DH, Valadares Filho SC, de Carvalho IPC, Martins JM. Níveis de proteína bruta em dietas para bovinos de corte: consumo, digestibilidade e desempenho produtivo. Rev Bras Zootec. 2006;35(6):2434-42.
29. Gleghorn JF, Elam NA, Galyean ML, Duff GC, Cole NA, Rivera JD. Effects of crude protein concentration and degradability on performance, carcass characteristics, and serum urea nitrogen concentrations in finishing beef steers. J Anim Sci. 2004;82(9):2705-17.
30. Galyean ML, Hubbert ME. Traditional and alternative sources of fiber: roughage values, effectiveness, and levels in starting and finishing diets. Prof Anim Sci. 2014;30:571-84.
31. Cole NA, Defoor PJ, Galyean ML, Duff GC, Gleghorn JF. Effects of phase-feeding of crude protein on performance, carcass characteristics, serum urea nitrogen concentration, and manure nitrogen of finishing beef steers. J Anim Sci. 2006;84(12):3421-32.
32. Waldrip HM, Todd RW, Cole NA. Prediction of nitrogen excretion by beef cattle: a meta-analysis. J Anim Sci. 2013;91(9):4290-302.
33. Di Marco ON, Aello MS. ¿Afecta el exceso de amonio ruminal el gasto energético de rumiantes?. Engormix [Internet]. 2006 May 11 [cited 2020 Sep 25]. Available from: https://bit.ly/36jWL0n.
34. Poppi DP, McLennant SR. Protein and energy utilization by ruminants at pasture. J Anim Sci. 1995;73(1):278-90.
35. Owens FN, Dubeski P, Hansont CF. Factors that alter the growth and development of ruminants. J Anim Sci. 1993;71(11):3138-50.
36. Bartlett KS, McKeith FK, VandeHaar MJ, Dahl GE, Drackley JK. Growth and body composition of dairy calves fed milk replacers containing different amounts of protein at two feeding rates. J Anim Sci. 2006;84(6):1454-67.
37. Donnelly PE, Hutton JB. Effects of dietary protein and energy on the growth of Friesian bull calves. N Z J Agric Res. 1976;19(4):409-14.
Author notes
beretta@fagro.edu.uy
Additional information
Authors contribution statement: VB:
Conceptualization, data analysis, article writing and editing; AS:
conceptualization, supervision, data collection, and manuscript review; IM:
data collection; IY: data collection.
Editor: The following editors approved this
article.
Cristina
Cabrera
ORCID: https://orcid.org/0000-0002-7964-6669 Universidad
de la República, Montevideo, Uruguay.
Alternative link
http://agrocienciauruguay.uy/ojs/index.php/agrociencia/article/view/111/413 (pdf)





