1. Introducción
La acidez y la escasa disponibilidad de nutrimentos son algunas de las razones de la baja productividad en los suelos (Baligar et al., 1998; Molina, 1998; Zapata, 2004). Se estima que los suelos ácidos ocupan alrededor de 3,95 billones de hectáreas, lo que corresponde a un 30% de la superficie cultivable del mundo (von Uexküll & Mutert, 1995). Se estima, además, que el 35% de las tierras cultivables de América Central son ácidas reflejando el grado de degradación, meteorización y lixiviación que han experimentado (Baligar et al., 1998). Sin embargo, la acidificación de las tierras dedicadas al cultivo del banano en Costa Rica es causada principalmente por la utilización de fertilizantes de reacción ácida (Molina, 1998; Serrano, 2003). Al establecerse originalmente las plantaciones se seleccionaron tierras sin acidez, debido a que ésta es una característica no deseable cuando se escogen tierras para la siembra del banano (Jaramillo et al., 1979; Jaramillo & Vázquez, 1900).
Los efectos adversos de la acidez sobre el crecimiento de los cultivos se deben a una serie de factores que incluyen bajo pH, toxicidad de Al y/o Mn y deficiencias de P, Ca y Mg (Ayarza & Salinas, 1982; Matsumoto, 2002). El Al y el Mn intercambiables son los principales elementos tóxicos en la mayoría de los suelos ácidos, el Al puede ocupar entre 4 y 94% de la capacidad de intercambio catiónico (Matsumoto, 2002). La toxicidad del Al es el factor más perjudicial para las plantas en los suelos ácidos, particularmente cuando el pH es inferior de 5,5 (Bertsch, 1995). El valor de pH en el cual los niveles del Al alcanzan efectos perjudiciales es diferente entre especies y también entre variedades o cultivares de una misma especie (Tang et al., 1993; Van et al., 1997; Zhang & Jessop, 1998), pero este valor depende también de ciertas condiciones del suelo como la mineralogía de las arcillas, el contenido de la materia orgánica, la presencia de otros cationes y aniones y de la salinidad total del suelo (Fassbender & Bornemisza, 1987).
Experimentos en soluciones nutritivas han demostrado que concentraciones de Al que fluctúan entre 0,2 a 6,0 mg l-1 son suficientes para limitar el crecimiento de varias especies de plantas (Ayarza & Salinas, 1982; Foy, 1984; Spain et al., 1975; Voight et al., 1999). Estos resultados se obtuvieron en experimentos con soluciones nutritivas y en suelo sobre agar en los cuales se añadieron varias concentraciones de Al. En general, los experimentos fueron diseñados para estimar el efecto de dosis de Al en períodos cortos. La reducción en la elongación de las raíces es el primer síntoma visible de la toxicidad del Al en muchas especies de plantas. El Al en concentraciones de micromoles en la solución inhibe fuertemente la división celular en las raíces y, en consecuencia, su crecimiento. Sin embargo, la tasa de reducción de la elongación disminuye en presencia de otros iones como el Ca y Mg, debido a que la interacción de estos cationes con el Al reduce la toxicidad. La inhibición de la elongación, así como de la división de las células en la raíz, son consecuencia de la interacción del Al con las moléculas de ADN, lo que cambia la fluidez y la permeabilidad de las membranas celulares, reduce la actividad de la ATPasa ligada a las membranas, inhibe la absorción de Ca, Mg y precipita los fosfatos (Cronan & Grigal, 1995). Cambios de carga eléctrica en la superficie de la raíz y la presencia de otros iones, especialmente cationes, disminuyen la absorción del Al (Alva et al., 1986; Ayarza & Salinas, 1982; Foy, 1984; Matsumoto, 2002; Ritchey et al., 1988). El Al absorbido por las plantas tiende a acumularse más en las raíces que en la parte aérea, variando la concentración según la especie. Se ha tratado de asociar la tolerancia al Al+3 con la habilidad de las plantas para acumular o excluir el Al en la parte aérea, pero los resultados han sido contradictorios, lo que indica que la tolerancia es una característica propia de cada especie (Foy, 1984).
El uso de fertilizantes de reacción neta ácida (úrea, nitrato de amonio, sulfato de amonio, fosfato monoamónico y fosfato diamónico) contribuye a la acidificación del suelo de manera proporcional al índice de acidez fisiológica del fertilizante (Kass, 1996; Tisdale et al., 1993). El grado de acidificación de los suelos bananeros depende también del manejo y la edad de la plantación, en forma general, a mayor edad de la plantación mayor es la acidificación de los suelos. Se ha demostrado que el uso de fertilizantes de reacción ácida y la lixiviación de bases en el agua de drenaje de un suelo cultivado con banano durante un período de dos año redujeron el pH de 6,48 a 5,07 e incrementaron la acidez intercambiable de 0,07 a 1,16 cmol(+) l-1 (Serrano, 2003). En Costa Rica existen 43.031 has cultivadas con banano (CORBANA, 2011) y es de esperarse que al menos el 70 % de las tierras dedicadas al cultivo presenten algún grado de acidificación.
Considerando que las musáceas son de los principales cultivos de las zonas tropicales, existen pocos estudios acerca de la sensibilidad de estas plantas a la toxicidad del Al (Delvaux, 1995; Sharrock & Frison, 1999; Wilson et al., 1987). La acidez reduce entre un 10 a 50% la cantidad de masa radicular en la planta de banano, dependiendo del nivel de acidificación de los suelos, en consecuencia, la productividad se puede reducir entre 50 a 67%, si la acidez es el único factor limitante (Gaugel et al., 2003). Se ha observado que las plantas de banano acumulan Al en las raíces y cormo, mientras que la acumulación en la parte aérea es relativamente baja, lo que sugiere que la planta de banano no es tolerante al Al (Gaugel et al., 2003; Rufyikiri et al., 2000, 2001). Por otro lado, los plátanos pueden tolerar hasta un 77% de saturación de Al en el complejo de intercambio (Rodríguez-García et al., 1985). Investigación con varios cultivares de plátanos y un cultivar de banano (Gran Naine) demostró que una concentración de 78,5 uM de Al redujo la absorción de agua entre 30 y 40 % con respecto al testigo. La presencia de Al redujo también la absorción de Ca, Mg, K, N-NO3 y N-NH4 y especialmente la de Mg que disminuyó en 80% (Rufyikiri et al., 2000, 2001).
El incremento en la concentración de Al en el medio disminuye la absorción del Mg y el contenido de este nutrimento en diferentes partes de la planta. El efecto del Al en el crecimiento de la planta se puede prevenir con la presencia de altos niveles de Mg (alta relación Mg/Al) evitando así las deficiencias de Mg inducidas por la presencia del Al (Marschner, 1995). El Mg se encuentra en mayor proporción en las vacuolas, aunque también se encuentra abundantemente en el citosol donde ayuda a mantener el pH entre 6,5 y 7,5, mientras que los cloroplastos contienen entre 10 y 20% del total de Mg en la hoja (Gil, 1995). Entonces, el incremento del Mg en la solución podría llegar a compensar los efectos tóxicos del Al en las plantas de banano (Gil, 1995; Marschner, 1995; Rufyikiri et al., 2001).
El objetivo del presente trabajo fue evaluar el efecto del pH y el Mg en la toxicidad del Al y en el crecimiento y absorción de nutrimentos y agua de plantas jóvenes de banano creciendo en una solución nutritiva.
2. Materiales y métodos
El experimento se condujo en el Centro de Investigación San Luis de la Compañía Bananera Atlántica (CISL-COBAL), en San Luis, distrito primero del cantón de Guácimo provincia de Limón, Costa Rica. El CISL está ubicado a 82 msnm, tiene una temperatura promedio anual de 27 ºC y una precipitación anual promedio de 3.970 mm. Se utilizaron plantas provenientes de cultivo de tejidos del clon “Gran Naine” de 7 a 10 cm de altura y con 4 a 5 hojas emitidas que tenían aproximadamente 4 semanas de edad y que estaban listas para iniciar la etapa de “endurecimiento”. Las plantas se pusieron a crecer en potes de 20.0 l de capacidad que contenían la solución nutritiva propuesta por Johnson y descrita por Lorenz & Maynard (1988). El volumen de solución nutritiva en cada pote fue de 19.0 l. La concentración de nutrimentos (mg l-1) de la solución base fue: N: 104; P: 33; K: 138; Mg: 25; Ca 90; S: 35; Fe: 2,6; Cu: 0,1; Zn: 0,3; Mn 1; B 0,3 y Mo: 0,01 y se la cambió dos veces por semana. Se evaluaron los siguientes tratamientos: 1) 25 mg l-1 de Mg + 0 mg l-1 de Al, pH 4; 2) 25 mg l-1 de Mg + 25 mg l-1 de Al, pH 3; 3) 50 mg l-1 de Mg + 25 mg.l-1 de Al, pH 3; 4) 100 mg l-1 de Mg + 25 mg l-1 de Al, pH 3; 5) 200 mg l-1 de Mg + 25 mg l-1 de Al, pH 3, y 6) 25 mg l-1 de Mg + 25 mg l-1 de Al, pH 5. Los tratamientos 1 y 2 sirvieron para evaluar el efecto del Al+3 a una misma concentración de Mg, mientras que los tratamientos del 2 al 5 permitieron evaluar el efecto de las dosis de Mg sobre el Al+3 y los tratamientos 2 y 6 sirvieron para medir el efecto del pH sobre la toxicidad del Al+3. Para suplir el Mg adicional al suministrado en la solución nutritiva base (tratamientos 3, 4 y 5) se preparó una solución madre de 23.750 mg l-1 de Mg con Mg- SO4.7H2O y se tomaron alícuotas de 20, 60 y 140 ml por cada pote con 19 l de agua. El aporte de Al se hizo utilizando Al2(SO4)3.18H O, preparando una solución madre de 9.500 mg l-l y tomando alícuotas de 50 ml por cada pote con 19 l de agua. La concentración del S (mg l-1) fue diferente en los distintos tratamientos por efecto del ión acompañante de las fuentes usadas para aportar el Mg y el Al, así 2 (79), 3 (112), 4 (177), 5 (307), 6 (79) con respecto a la indicada anteriormente en la solución base que fue la utilizada en el tratamiento 1 (35). Para obtener pH 5 fue necesario sustituir el KNO3 de la solución Johnson por K2CO3 y NH4NO3 con la preparación de soluciones madre de 91.463 mg l-1 de K y de 31 688 mg l-1 de N y se tomaron alícuotas de 20 ml por pote. Se utilizó un diseño irrestricto al azar con 5 repeticiones, cada planta constituyó la unidad experimental respectiva. Al inicio y a las 9 semanas de iniciado el experimento se evaluaron las siguientes variables: 1) ritmo de emisión foliar (REF) utilizando la siguiente fórmula: N° de hojas totales emitidas / N° de semanas evaluadas. Al momento de establecer el experimento se marcó la última hoja emitida completamente abierta y se tomó nota del estado de la hoja “candela”, a partir de ahí se contaron las hojas emitidas al final de las 9 semanas; 2) índice de incremento en altura (IIA) que se logró midiendo desde la base del cormo hasta la “V” formada entre la última hoja completamente abierta y la hoja candela. El IIA se obtuvo a partir de la fórmula (cm de altura final – cm altura inicial) / N° de semanas evaluadas; 3) área foliar (AF) medida al inicio y final del experimento solamente en las hojas completamente abiertas. Se procedió a dibujar las hojas en papel, luego se recortó el dibujo por el perímetro y se determinó la masa del papel. El área se obtuvo al comparar la masa del dibujo de la hoja contra la masa de un dibujo del mismo tipo de papel de área conocida (400 cm2); 4) producción de biomasa (PB) determinada a las 9 semanas de iniciados los tratamientos. Se dividió la planta en raíz de segundo orden, raíz de primer orden, cormo, pseudo tallo y nervadura (incluye pseudo peciolo) de hojas inferiores y superiores; 5) lámina foliar (LF) de las hojas inferiores y superiores. La división de hojas superiores e inferiores fue arbitraria y se consideró el grupo de hojas superiores a las últimas 5 hojas emitidas (hojas más jóvenes) y el grupo de hojas inferiores a las restantes hojas al momento de la cosecha. Todos estos tejidos se secaron en estufa a 70 °C hasta alcanzar masa constante. Se calcularon los siguientes índices fisiológicos y parámetros morfológicos de acuerdo a las fórmulas respectivas descritas por Bertsch (1995), Rodríguez & Leihner (2006) y Turner (1972): 1) índice de crecimiento absoluto (ICA) = (msf – msi)/t2 – t1)) donde msf=masa seca final, msi= masa seca inicial, t2=tiempo final, t1= tiempo inicial; 2) índice de crecimiento relativo (ICR) = ((ln msf-ln msi)/ (t2-t1)), (ln=logaritmo natural); 3) índice de crecimiento foliar absoluto (ICFA): ((af2 - af1)/(t2-t1)), (af2 = área foliar final, af1 = área foliar inicial); 4) índice de crecimiento foliar relativo (ICFR): ((ln af2-ln af1)/ (t2-t1)); 5) tasa de asimilación neta (TAN): (((msf-msi)x(ln af2 – ln af1))/(t2-t1)x(af2-af1); 6) relación de área foliar (RAF): (af2 / msf); 6) relación de masa foliar (RMF): (msfo/msf), (msfo = masa seca foliar); 7) área foliar específica (AFE): (af2/msfo); 7) razón de masa radicular (RMR): (msr/msf), (msr = masa seca de raíces); relación de raíces a hojas (RRH): (msr/mofo) y; relación de raíces a parte aérea (RRPA): (msr/ mspa), (mspa= masa seca parte aérea).
Debido a que el método de determinación de la biomasa fue destructivo, la masa seca inicial se determinó de manera indirecta, tomando 10 plantas de iguales características a las utilizadas en el experimento en el mismo momento del establecimiento y se determinó la masa fresca y seca para calcular el porcentaje de masa seca por planta. En cada planta se determinó el peso fresco antes de colocarse en el pote respectivo y se asumió que las mismas presentaban igual porcentaje de masa seca que el promedio de las 10 plantas utilizadas en la determinación del porcentaje de masa seca, obteniendo así la masa seca inicial de cada una de las plantas usadas en cada tratamiento. Al momento de la cosecha se determinó la concentración de nutrimentos en los diferentes órganos de la planta. El análisis químico se realizó por el método de digestión húmeda (Westerman, 1990), el material seco de cada tejido de la planta fue molido y se hizo una digestión con ácido nítrico y se determinaron los contenidos de P, K, Ca, Mg, S, Fe, Cu, Zn, Mn, y B por espectrometría de emisión de plasma, el N se determinó por el método de combustión en seco (Dumas) en analizador automático de N (Westerman, 1990). El análisis de la conductividad eléctrica se realizó por el método del puente eléctrico, utilizándose un conductímetro marca Beckman Instruments, Inc. Modelo RC-16C, el cual fue calibrado con una solución KCl 0,01M (Westerman, 1990). La absorción total de nutrimentos se calculó multiplicando la concentración de cada nutrimento por la masa seca de los diferentes componentes de la biomasa y se realizó la sumatoria para obtener la absorción total.
El consumo de agua se determinó aforando los potes a 19 litros con solución nutritiva, se determinó la masa del pote + solución nutritiva al inicio y final de cada ciclo de sustitución (3 y 4 días) y la diferencia de masa correspondió al consumo de agua en litros por la planta en dicho período. El consumo total de agua se obtuvo a partir de la sumatoria de agua consumida en cada ciclo de sustitución de la solución nutritiva. El pH de la solución se midió con el método del potenciómetro (Westerman, 1990) y se controló cada 24 horas y se ajustó siempre que la lectura presentara una desviación de -0,17 y + 0,3 de unidad de pH mediante la adición de KOH 0,5M y HCl concentrado, según fuera el caso.
Para el análisis estadístico se utilizó el modelo lineal general (GLM) procedimiento del programa S.A.S versión 8.1 (análisis de varianza) y pruebas de contraste. Se consideraron las diferencias significativas a valores de p ≤ 0,05, la separación de medias se realizó por la prueba de diferencia mínima significativa.
3. Resultados y discusión
3.1. Consumo total de agua
El consumo de agua por las plantas de banano disminuyó significativamente con la aplicación de Al (T1 vs T2) (Figura 1). Resultados similares han sido reportados en otras especies (Wagatsuma 1983, Keltjens 1983, Tice 1992, Ishikawa & Wagatsuma 1998, Matsumoto 2002). El menor consumo de agua por las plantas tratadas con Al fue el primer síntoma observado y las diferencias fueron estadísticamente significativas a partir del tercer día. Investigación conducida por Rufyikiri et al. (2001) encontró también que el primer síntoma que presentaron las plantas de banano y plátano tratadas con Al fue la disminución en el consumo de agua, las diferencias en banano Gran Naine fueron significativas a partir del día catorce de aplicado el Al. Es probable que las diferencias en el tiempo de detección del síntoma se deban a que las concentraciones de Al en la solución utilizada por Rufykiri y colaboradores fue menor por lo que la reducción en consumo de agua tardó más tiempo. En las plantas tratadas con Al no se encontraron cambios en el consumo de agua al incrementar las dosis de Mg hasta el día 38, momento que coincidió con el crecimiento más acelerado de las plantas. Un mayor consumo de agua sólo se presentó con la dosis de 50 mg l-1 de Mg, consumo que descendió en las dosis superiores, debido al aumento de conductividad eléctrica de las dosis más altas de Mg (Figura 2) que ocasionó un menor consumo de agua por parte de la planta (De Oliveira et al., 2000). El incremento de pH de 3 a 5 fue más efectivo para mejorar la absorción de agua que el aumento en la dosis de Mg (Figura 1).
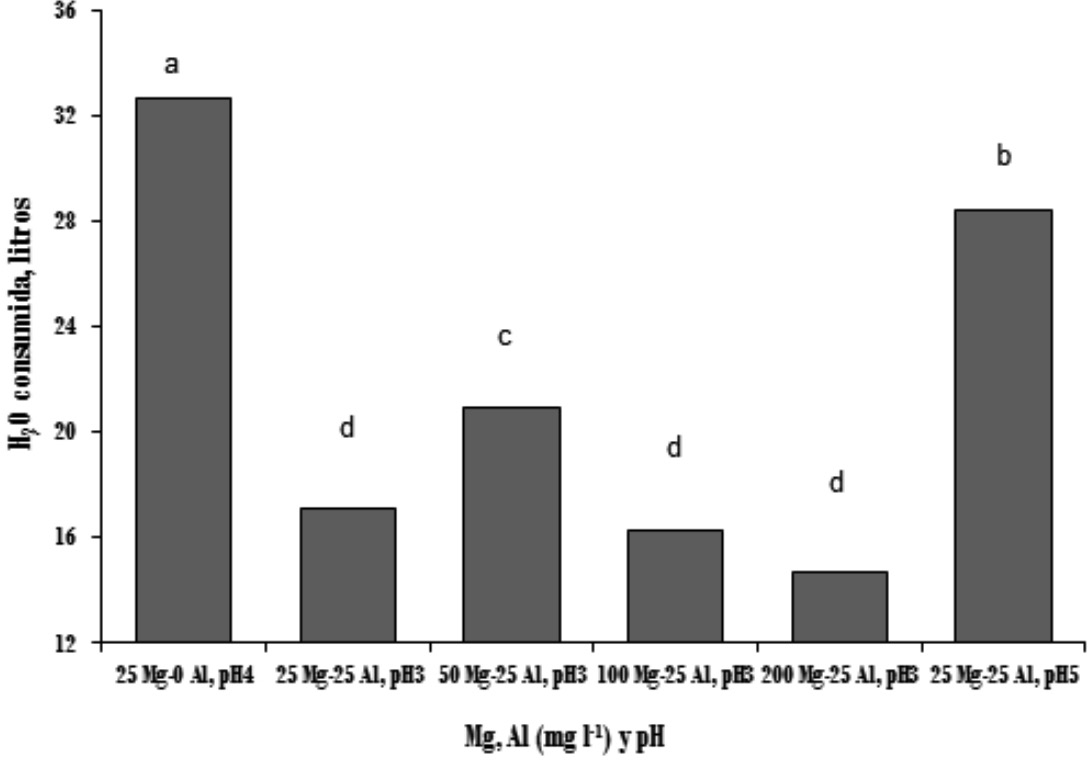 Figura 1
Consumo
acumulado de agua a los 63 días de iniciado el experimento.
Figura 1
Consumo
acumulado de agua a los 63 días de iniciado el experimento.
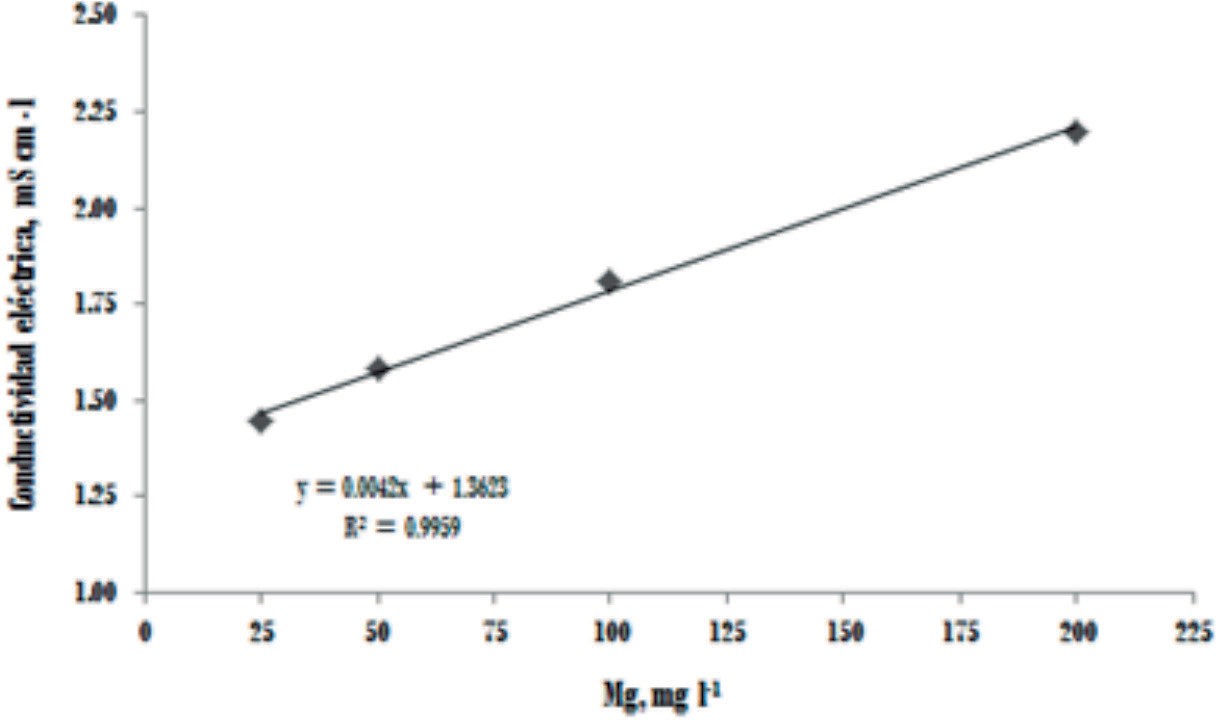 Figura 2
Conductividad
eléctrica en función de la concentración de Mg en la solución nutritiva.
Figura 2
Conductividad
eléctrica en función de la concentración de Mg en la solución nutritiva.
3.2. Descripción
de síntomas de toxicidad de Al en plantas de banano
Los principales
síntomas observados en las plantas creciendo en la solución nutritiva con Al
con la menor dosis de Mg (25 mg l-1) fueron los siguientes: clorosis
marcada a nivel de las hojas inferiores y reducción en el distanciamiento entre
los pseudo pecíolos (Figura 3a). Como consecuencia de lo anterior, las plantas
se tornaron más compactas (tipo roseta) y se observó una obstrucción foliar
(arrepollamiento), lo que a su vez provocó una
reducción de la altura en las
plantas. De igual forma,
las hojas de las plantas creciendo en solución
nutritiva con Al fueron de menor tamaño y el pseudo tallo tuvo menor circunferencia. La reducción
del tamaño de la hoja y de
la distancia entre pseudo pecíolos es una respuesta del
banano al déficit de agua
(Turner, 1972). Síntomas similares fueron
observados por Rufyikiri et al. (2001) quienes
sugirieron que estos efectos pueden resultar de la reducción
de crecimiento y de actividad radicular
inducidos por la presencia de Al, condición que hace que la planta
absorba menos agua.
 Figura 3
Efecto
de la aplicación de Al en plantas de banano Gran Naine: (a) efecto de la
toxicidad del Al en la reducción del distanciamiento entre pseudo peciolos; (b)
inicio de clorosis; (c) necrosis avanzando hacia nervadura central; (d) efecto
de concentración de Mg en la solución sobre la presencia de la clorosis.
Figura 3
Efecto
de la aplicación de Al en plantas de banano Gran Naine: (a) efecto de la
toxicidad del Al en la reducción del distanciamiento entre pseudo peciolos; (b)
inicio de clorosis; (c) necrosis avanzando hacia nervadura central; (d) efecto
de concentración de Mg en la solución sobre la presencia de la clorosis.
La clorosis se
inició en los bordes de las hojas inferiores y fue ascendiendo a las hojas
superiores (Figura 3b) avanzando luego hacia la nervadura central, pero ésta
permaneció verde (Figura 3c), la lámina foliar se decoloró y se intensificó la
clorosis con el paso del tiempo hasta que los tejidos se necrosaron. Esta
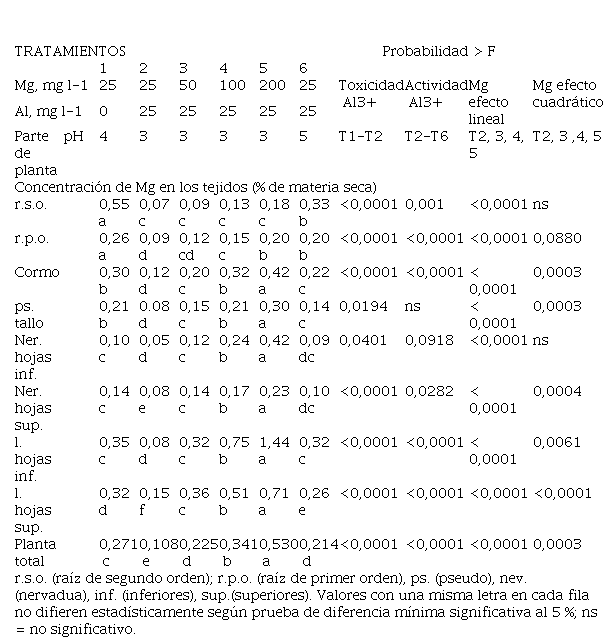
sintomatología aparece por deficiencia de Mg (Tabla 1). El Mg es muy móvil
en la planta, por esta razón la clorosis
es ascendente (Villalobos & Killorn, 2001). La clorosis en las hojas
inferiores solo se presentó en las plantas que crecieron en la solución con la dosis menor de Mg
a la que se aplicó Al (Figura 3d) lo que confirma el efecto positivo de la
adición de cantidades mayores de Mg para evitar que se desarrolle la clorosis
propia de los síntomas de carencia de Mg. Estos resultados demostraron que la
clorosis fue inducida por la presencia del Al a través de una interferencia
significativa de la absorción del Mg (Tabla 1). Los mismos síntomas fueron
documentados por Rufyikiri et al. (2000) al evaluar el efecto del
Al en plantas de banano y otros autores (Martin-prevel, 1980; Turner, 1972) han
asociado también esta sintomatología a la deficiencia de Mg.
Tabla 1
Efecto del pH, la concentración del Mg
en la solución nutritiva y el Al sobre la concentración de Mg en las diferentes
partes de la planta analizadas.
|
TRATAMIENTOS Probabilidad > F
|
| |
1
|
2
|
3
|
4
|
5
|
6
| | | | |
|
Mg, mg l-1
|
25
|
25
|
50
|
100
|
200
|
25
|
Toxicidad Al3+
|
Actividad Al3+
|
Mg efecto lineal
|
Mg efecto cuadrático
|
|
Al, mg l-1
|
0
|
25
|
25
|
25
|
25
|
25
|
|
Parte
de planta
|
pH
|
4
|
3
|
3
|
3
|
3
|
5
|
T1-T2
|
T2-T6
|
T2, 3, 4, 5
|
T2, 3 ,4, 5
|
|
Concentración
de Mg en los tejidos (% de materia seca)
|
|
r.s.o.
| |
0,55 a
|
0,07 c
|
0,09 c
|
0,13 c
|
0,18 c
|
0,33 b
|
<0,0001
|
0,001
|
<0,0001
|
ns
|
|
r.p.o.
| |
0,26 a
|
0,09 d
|
0,12 cd
|
0,15 c
|
0,20 b
|
0,20 b
|
<0,0001
|
<0,0001
|
<0,0001
|
0,0880
|
|
Cormo
| |
0,30 b
|
0,12 d
|
0,20 c
|
0,32 b
|
0,42 a
|
0,22
c
|
<0,0001
|
<0,0001
|
< 0,0001
|
0,0003
|
|
ps. tallo
| |
0,21 b
|
0.08
d
|
0,15 c
|
0,21 b
|
0,30 a
|
0,14
c
|
0,0194
|
ns
|
< 0,0001
|
0,0003
|
|
Ner. hojas inf.
| |
0,10 c
|
0,05 d
|
0,12 c
|
0,24 b
|
0,42 a
|
0,09 dc
|
0,0401
|
0,0918
|
<0,0001
|
ns
|
|
Ner.
hojas sup.
| |
0,14 c
|
0,08 e
|
0,14 c
|
0,17 b
|
0,23 a
|
0,10 dc
|
<0,0001
|
0,0282
|
< 0,0001
|
0,0004
|
|
l. hojas inf.
| |
0,35 c
|
0,08 d
|
0,32 c
|
0,75 b
|
1,44 a
|
0,32 c
|
<0,0001
|
<0,0001
|
< 0,0001
|
0,0061
|
|
l.
hojas sup.
| |
0,32 d
|
0,15 f
|
0,36 c
|
0,51 b
|
0,71 a
|
0,26 e
|
<0,0001
|
<0,0001
|
<0,0001
|
<0,0001
|
|
Planta total
| |
0,271 c
|
0,108 e
|
0,225 d
|
0,341 b
|
0,530 a
|
0,214
d
|
<0,0001
|
<0,0001
|
<0,0001
|
0,0003
|
|
r.s.o. (raíz de segundo
orden); r.p.o. (raíz de primer orden), ps. (pseudo), nev. (nervadua),
inf. (inferiores), sup.(superiores). Valores con una misma letra en cada fila no difieren estadísticamente
según prueba de diferencia mínima
significativa al 5 %; ns = no significativo.
|
La concentración de Mg en las raíces disminuyó en presencia de Al, aún en las dosis más altas de Mg (Tabla 1). La concentración de Mg en el cormo y pseudo tallo también disminuyó con la presencia del Al en el medio de crecimiento, pero se recuperó a partir de la dosis de 100 mg l-1 de Mg. En el caso de las hojas y la planta total, el Al redujo la concentración de Mg en estos tejidos de forma significativa solo con la dosis más baja de Mg (25 mg l-1), al subir la dosis de Mg se produjo una recuperación de la concentración de Mg en esos tejidos. Los valores de concentración del Mg a nivel foliar fueron normales y adecuados en los tratamientos 1(Mg 25 mg l-1, Al 0 mg l-1 y pH 4), 3 (Mg 50 mg l-1, Al 25 mg l-1 y pH 3) y 6 (Mg 25mg l-1, Al 25 mg l-1 y pH 5), altos en los tratamientos 4 (Mg 100 mg l-1, Al 25 mg l-1 y pH 3) y 5 (Mg 200 mg l-1, Al 25 mg l-1 y pH 3) y bajos en el tratamiento 2 (Mg 25 mg l-1, Al 25 mg l-1 y pH 3).
3.3. Efecto de la concentración de Mg y del pH en las variables de crecimiento
La aplicación de Al afectó en forma significativa las variables de crecimiento como acumulación de biomasa, altura de la planta, área foliar, circunferencia y número de hijos emitidos. El Al no tuvo efecto significativo en el número de hojas emitidas, ritmo de emisión foliar y el número de hojas al momento de finalizar el experimento (Tabla 2) coincidiendo con los resultados encontrados por Rufyikiri et al. (2000). Se ha demostrado que la emisión foliar y la tasa de crecimiento del fruto están gobernados principalmente por la temperatura ambiental (Robinson, 1996; Stover & Simmonds, 1987; Turner & Lahav, 1983).
Por otro lado, se observó un efecto favorable al incrementar el Mg de 25 a 50 mg l-1 en la altura de la planta y la circunferencia en la base del pseudo tallo, sin embargo, este incremento no fue suficiente para neutralizar el efecto del Al. Dosis superiores a 50 mg l-1 no tuvieron ningún efecto sobre la altura de la planta y la circunferencia del pseudo tallo debido al incremento en la conductividad eléctrica de la solución nutritiva al elevar la concentración de Mg (Figura 2), condición que afectó en forma negativa el crecimiento de la planta. La utilización de dosis mayores de 50 mg l-1 redujo el efecto positivo del incremento de Mg en la solución, lo que confirma que el banano es una planta susceptible a la salinidad (De Oliveira et al., 2000).
El incremento del pH de 3
a 5 fue más efectivo en disminuir la toxicidad del Al en las variables de
crecimiento de la planta que el incremento de la dosis de Mg en la solución
nutritiva (Tabla 2). Lo anterior se podría explicar por la presencia de las diferentes especies monoméricas de Al a diferente pH, especialmente el
Al+3 y el Al(OH)+2. Generalmente,
el Al+3 es más fitotóxico que el Al(OH)+2 y esta especie
es más tóxica que el Al(OH)2+, a pH 4 el Al está presente como Al+3, mientras que a pH 5 coexisten en equilibrio Al(OH)2+ y Al+3 (Miyasaka et al., 2007). Se considera
que las dicotiledóneas son más sensitivas al Al(OH)2+ que al Al+3,
mientras que para las monocotiledóneas
lo son al Al+3 posiblemente debido a
que las monocotiledóneas tienen mayor capacidad de intercambio catiónico en la pared celular (Alva et al.,
1986). Los resultados de este estudio sugieren que la planta de banano es afectada principalmente por Al+3 cuando el pH es igual o
menor a 4,0.
Tabla 2
Efecto de la
concentración del Mg y pH en la actividad del Al en las variables de
crecimiento evaluadas.

|
TRATAMIENTOS Probabilidad > F
|
| |
1
|
2
|
3
|
4
|
5
|
6
| | | | |
|
Mg, mg l-1
|
25
|
25
|
50
|
100
|
200
|
25
|
Toxicidad
Al3+
|
Actividad
Al3+
|
Mg efecto lineal
|
Mg efecto
cuadrático
|
|
Al, mg l-1
|
0
|
25
|
25
|
25
|
25
|
25
| | |
|
Variables
|
pH
|
4
|
3
|
3
|
3
|
3
|
5
|
T1-T2
|
T2-T6
|
T2, 3, 4, 5
|
T2, 3 ,4, 5
|
| m.s.f
(g)
| | 153,91 a
| 53,13 c
| 67,28 c
| 54,71
c
| 52,72 c
| 123,00 b
| <0,0001
| <0,0001
| ns
| ns
|
|
a.f.
(cm)
| | 74,40 a
| 41,33 d
| 49,63 c
| 42,96
cd
| 40,30 d
| 60,60 b
| <0,0001
| <0,0001
| ns
| ns
|
|
r.e.f.
(h sem-1)
| | 1,10 a
| 11,1 a
| 1,09 a
| 1,10 a
| 1,08 a
| 1,15 a
| ns
| ns
| ns
| ns
|
|
n.h.c.
| |
9,44 a
|
9,15 a
|
9,00 a
|
9,32
a
|
9,52 a
|
9,70 a
|
ns
|
ns
|
ns
|
ns
|
|
Ar. Fol.
(cm2)
| | 13.912 a
| 5.725 c
| 7.143
c
| 5.850
c
| 5.621 c
| 10.440 b
| <0,0001
| ns
| ns
| ns
|
|
c.b.
pt (cm)
| | 23,70 a
| 13,97 d
| 16,38 c
| 15,14
cd
| 14,80 cd
| 21,28 b
| 1<0,0001
| <0,0001
| ns
| ns
|
|
n.b.e.
“hijos”
| | 2,60 a
| 0,00 h
| 0,75 b
| 0,60 b
| 0,60 b
| 3,25 a
| <0,0001
| <0,0001
| ns
| 0,0080
|
|
m.s.f (masa seca final); a.f (altura final); r.e.f. (ritmo de emisión foliar, hojas por semana); n.h.c (número de hojas a cosecha); c.b. p.t. (circunferencia base del pseudo tallo); n.b.e (número de brotes emitidos). Valores con
una misma letra en cada fila no difieren estadísticamente según prueba de diferencia mínima significativa al 5 %; ns = no significativo.
|
3.4. Efecto de la concentración de
Mg y del pH en los índices de crecimiento y parámetros morfológicos
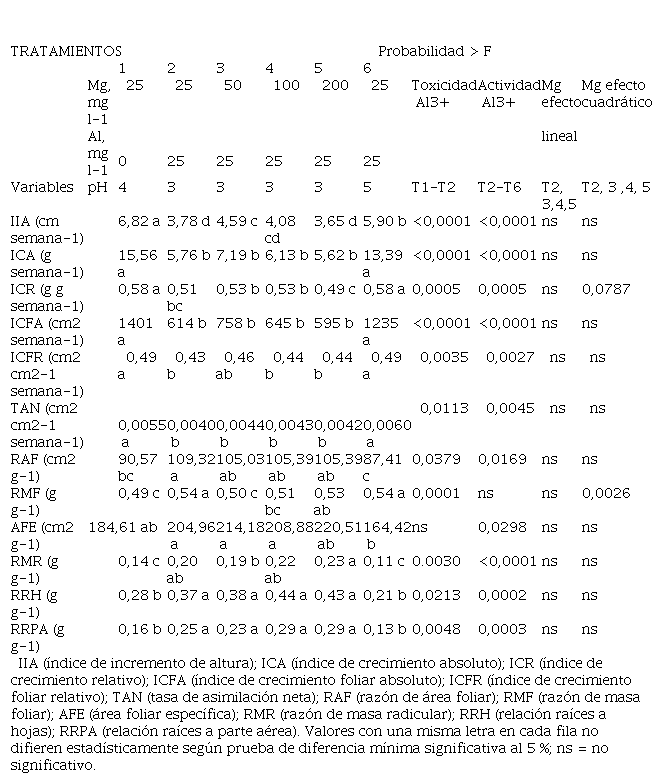
En general, el comportamiento de los índices de crecimiento estudiados fue la misma a la observada con los valores absolutos de las variables de crecimiento. El Al afectó de manera negativa los índices fisiológicos IIA, ICA, ICR, ICFA, ICFR y TAN (Tabla 3). La dosis de 50 mg l-1 de Mg disminuyó los síntomas de toxicidad, pero solo hubo diferencias significativas entre tratamientos con el IIA. Las dosis más altas de Mg no logran mayor efecto compensatorio sobre la toxicidad del Al, probablemente debido al efecto tóxico de la salinidad de la solución que fue causado por el incremento en las dosis de Mg.
La magnitud de los parámetros RAF, RMF, RMR, RRH, RRPA se incrementó con la aplicación de Al (Tabla 3), a pesar de que con estos tratamientos se redujo el área foliar, la masa seca total (Tabla 2) y la masa seca en las diferentes partes en que se dividió la planta (Tabla 4), por esta razón, las relaciones calculadas tienden a aumentar a menor biomasa producida en las diferentes partes de la planta. No se encontraron diferencias entre las dosis de Mg evaluadas en los valores de RAF, RRH, RRPA, pero sí con respecto al tratamiento sin Al y con Al a pH 5 que fueron los que mayor biomasa produjeron.
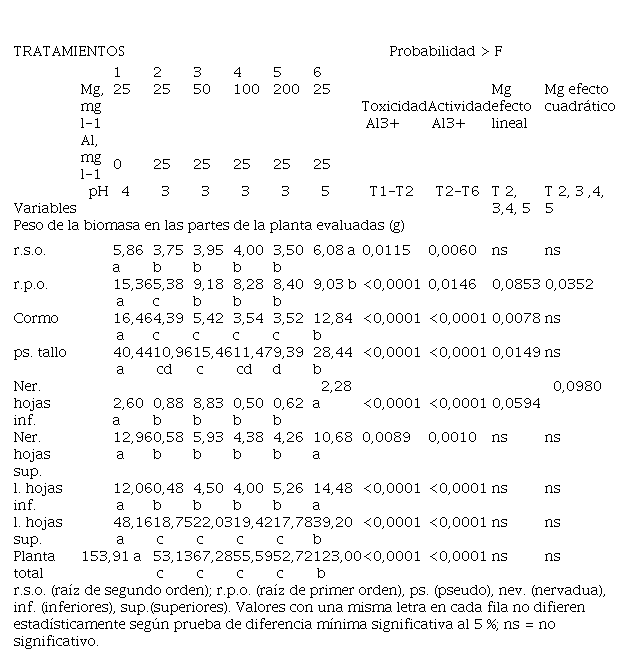
3.5. Efecto de la concentración de Mg y del pH en la producción de biomasa en las diferentes partes de la planta
La presencia
de Al en la solución afectó en forma negativa el desarrollo de las raíces, cormo, pseudo tallo, láminas foliares y nervaduras de hojas
(Tabla 4). Hubo
diferencias significativas con respecto al
tratamiento sin Al. Las raíces de primer orden
(primarias o raíces gruesas) fueron las más afectadas
por el Al. El incremento en la dosis de Mg disminuyó levemente los efectos del Al
en las raíces de primer orden
produciendo mayor cantidad de biomasa, lo mismo ocurrió en el tratamiento con Al con la menor dosis de Mg, pero a pH 5.
Tabla 3
Efecto de la
concentración del Mg y pH en la actividad del Al en los índices fisiológicos y
parámetros morfológicos evaluados.
|
TRATAMIENTOS Probabilidad > F
|
| |
1
|
2
|
3
|
4
|
5
|
6
| | | | |
|
Mg,
mg l-1 | 25
| 25
| 50
| 100
| 200
| 25
|
Toxicidad
Al3+
|
Actividad
Al3+
|
Mg efecto
lineal
|
Mg
efecto
cuadrático |
|
Al,
mg l-1 | | | | | | | | |
|
0
|
25
|
25
|
25
|
25
|
25
| | | | |
|
Variables |
pH
|
4
|
3
|
3
|
3
|
3
|
5
|
T1-T2
|
T2-T6
|
T2, 3,4,5
|
T2, 3 ,4, 5
|
|
IIA (cm
semana-1)
| |
6,82 a
|
3,78 d
|
4,59 c
|
4,08 cd
|
3,65 d
|
5,90 b
|
<0,0001
|
<0,0001
|
ns
|
ns
|
|
ICA (g semana-1)
| |
15,56 a
|
5,76 b
|
7,19
b
|
6,13 b
|
5,62 b
|
13,39 a
|
<0,0001
|
<0,0001
|
ns
|
ns
|
|
ICR (g g
semana-1)
| |
0,58 a
|
0,51 bc
|
0,53
b
|
0,53 b
|
0,49 c
|
0,58 a
|
0,0005
|
0,0005
|
ns
|
0,0787
|
|
ICFA (cm2 semana-1)
| |
1401 a
|
614 b
|
758 b
|
645 b
|
595 b
|
1235 a
|
<0,0001
|
<0,0001
|
ns
|
ns
|
|
ICFR (cm2 cm2-1 semana-1)
| | 0,49 a
| 0,43 b
| 0,46
ab
| 0,44 b
| 0,44 b
| 0,49 a
|
0,0035
| 0,0027
| ns
| ns
|
|
TAN (cm2
cm2-1 semana-1)
| | 0,0055 a
| 0,0040 b
| 0,0044 b
| 0,0043 b
| 0,0042 b
| 0,0060 a
|
0,0113
| 0,0045
| ns
| ns
|
|
RAF (cm2 g-1)
| |
90,57 bc
|
109,32 a
|
105,03 ab
|
105,39 ab
|
105,39 ab
|
87,41 c
|
0,0379
|
0,0169
|
ns
|
ns
|
|
RMF (g g-1)
| |
0,49 c
|
0,54 a
|
0,50
c
|
0,51 bc
|
0,53 ab
|
0,54 a
|
0,0001
|
ns
|
ns
|
0,0026
|
|
AFE (cm2 g-1)
|
184,61
ab
|
204,96 a
|
214,18 a
|
208,88 a
|
220,51 ab
|
164,42 b
|
ns
|
0,0298
|
ns
|
ns
|
|
RMR (g g-1)
| |
0,14 c
|
0,20 ab
|
0,19
b
|
0,22 ab
|
0,23 a
|
0,11 c
|
0.0030
|
<0,0001
|
ns
|
ns
|
|
RRH (g g-1)
| |
0,28 b
|
0,37 a
|
0,38
a
|
0,44 a
|
0,43 a
|
0,21 b
|
0,0213
|
0,0002
|
ns
|
ns
|
|
RRPA (g g-1)
| |
0,16 b
|
0,25 a
|
0,23 a
|
0,29 a
|
0,29 a
|
0,13 b
|
0,0048
|
0,0003
|
ns
|
ns
|
| IIA (índice de incremento de altura); ICA (índice de crecimiento absoluto); ICR (índice de crecimiento relativo); ICFA (índice
de crecimiento foliar absoluto); ICFR (índice de
crecimiento foliar relativo); TAN (tasa
de asimilación neta);
RAF (razón de área foliar); RMF (razón de masa foliar); AFE (área foliar
específica); RMR (razón
de masa radicular); RRH (relación raíces a hojas);
RRPA (relación raíces
a parte aérea). Valores con una misma letra en cada
fila no difieren estadísticamente según prueba de diferencia mínima significativa al 5 %; ns = no significativo.
|
El incremento
de las dosis de Mg no mejoró la producción de materia seca en el cormo y demás partes aéreas
cuando se lo comparó
con el tratamiento sin Al, sin embargo, el tratamiento
con Al a pH 5 y con la dosis
menor de Mg sí redujo parcialmente los efectos tóxicos del Al, alcanzando valores significativamente superiores al tratamiento con las misma
dosis de Mg, pero a pH 3. El incremento del pH de
3 a 5 con idéntica cantidad
de Mg logró disminuir los efectos de
la toxicidad del Al a nivel
de biomasa producida en las raíces
de segundo orden, nervadura y láminas
inferiores, lo anterior
posiblemente debido a una menor cantidad de Al+3 en la solución nutritiva en el tratamiento con pH 5, lo que probablemente
redujo el efecto negativo del Al en
la producción de biomasa aérea.
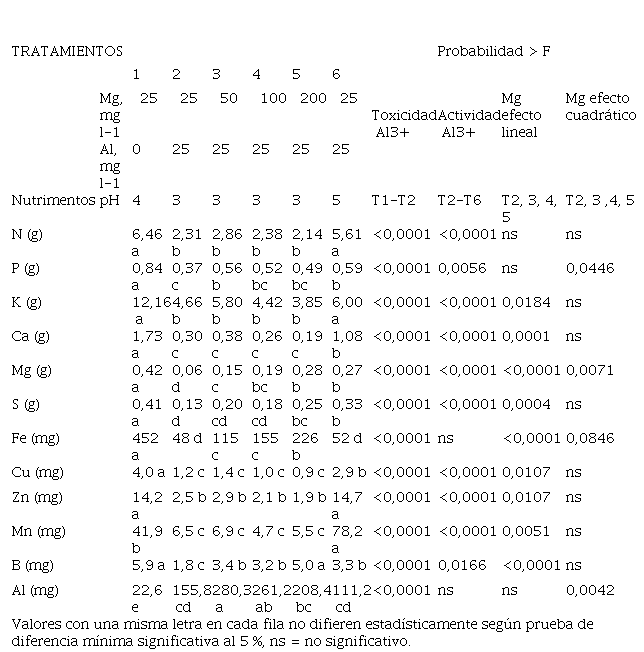
3.6. Efecto de la concentración de Mg y del pH en la absorción total de nutrimentos por la planta de banano
Se observó
que la aplicación de Al redujo la producción de biomasa (Tabla 4) lo cual hizo que la planta absorba solamente la tercera parte de N (Tabla 5). No
se encontró efecto de las dosis crecientes
de Mg sobre la producción de biomasa por parte
de la planta (Tabla 5), razón
por la cual no se afectó
la absorción de N. El incremento de pH de 3 a 5 mejoró significativamente la absorción de N, debido a que la concentración de Al+3 en la solución
es menor a pH 5 por la coexistencia de Al+3 con Al(OH)2+ en la solución (Miyasaka et al., 2007), lo
que reduce el efecto tóxico del Al. La absorción total del P
disminuyó significativamente por la aplicación de Al (Tabla 5), situación que se explica por la menor biomasa
producida por la planta cuando se adiciona Al (Tabla 4).
Tabla 4
Efecto de
la concentración del Mg y del pH en la
actividad del Al en la producción de biomasa en las diferentes partes de
la planta.
|
TRATAMIENTOS Probabilidad > F
|
| |
1
|
2
|
3
|
4
|
5
|
6
| | | | |
|
Mg, mg l-1
|
25
|
25
|
50
|
100
|
200
|
25
| Toxicidad
Al3+
| Actividad
Al3+
|
Mg efecto
lineal
|
Mg efecto
cuadrático
|
|
Al, mg l-1
| | | | | | |
|
0
|
25
|
25
|
25
|
25
|
25
| | | | |
| Variables
| pH
| 4
| 3
| 3
| 3
| 3
| 5
| T1-T2
| T2-T6
|
T
2, 3,4, 5
|
T
2, 3 ,4, 5
|
|
Peso de la biomasa
en las partes
de la planta evaluadas (g)
|
|
r.s.o.
| |
5,86 a
|
3,75 b
|
3,95 b
|
4,00 b
|
3,50 b
|
6,08 a
|
0,0115
|
0,0060
|
ns
|
ns
|
|
r.p.o.
| |
15,36 a
|
5,38 c
|
9,18 b
|
8,28 b
|
8,40 b
|
9,03 b
|
<0,0001
|
0,0146
|
0,0853
|
0,0352
|
|
Cormo
| |
16,46 a
|
4,39 c
|
5,42 c
|
3,54 c
|
3,52 c
|
12,84 b
|
<0,0001
|
<0,0001
|
0,0078
|
ns
|
|
ps. tallo
| |
40,44 a
|
10,96 cd
|
15,46 c
|
11,47 cd
|
9,39 d
|
28,44 b
|
<0,0001
|
<0,0001
|
0,0149
|
ns
|
|
Ner. hojas inf.
| | 2,60 a
| 0,88 b
| 8,83 b
| 0,50 b
| 0,62 b
| 2,28 a
| <0,0001
| <0,0001
| 0,0594
| 0,0980
|
|
Ner. hojas sup.
| |
12,96 a
|
0,58 b
|
5,93 b
|
4,38 b
|
4,26 b
|
10,68 a
|
0,0089
|
0,0010
|
ns
|
ns
|
|
l. hojas inf.
| |
12,06 a
|
0,48 b
|
4,50 b
|
4,00 b
|
5,26 b
|
14,48 a
|
<0,0001
|
<0,0001
|
ns
|
ns
|
|
l. hojas sup.
| |
48,16 a
|
18,75 c
|
22,03 c
|
19,42 c
|
17,78 c
|
39,20 b
|
<0,0001
|
<0,0001
|
ns
|
ns
|
|
Planta total
|
153,91 a
|
53,13 c
|
67,28 c
|
55,59 c
|
52,72 c
|
123,00 b
|
<0,0001
|
<0,0001
|
ns
|
ns
|
|
r.s.o. (raíz de segundo orden);
r.p.o. (raíz de primer orden), ps. (pseudo),
nev. (nervadua), inf. (inferiores),
sup.(superiores). Valores con una misma letra en cada
fila no difieren estadísticamente según prueba de diferencia mínima significativa al 5 %; ns = no significativo.
|
Tabla 5.
Efecto de la
concentración del Mg y pH en la actividad del Al en la absorción total de
nutrimentos por la planta de banano.
|
TRATAMIENTOS
| | | |
Probabilidad > F
| |
| |
1
|
2
|
3
|
4
|
5
|
6
| | | | |
|
Mg, mg l-1
| 25
| 25
| 50
| 100
| 200
| 25
| Toxicidad Al3+
| Actividad Al3+
|
Mg efecto lineal
|
Mg efecto
cuadrático
|
|
Al, mg l-1
|
0
|
25
|
25
|
25
|
25
|
25
| | | | |
|
Nutrimentos
|
pH
|
4
|
3
|
3
|
3
|
3
|
5
|
T1-T2
|
T2-T6
|
T2, 3, 4, 5
|
T2, 3 ,4, 5
|
|
N (g)
| |
6,46 a
|
2,31 b
|
2,86 b
|
2,38 b
|
2,14
b
|
5,61 a
|
<0,0001
|
<0,0001
|
ns
|
ns
|
|
P (g)
| |
0,84 a
|
0,37 c
|
0,56 b
|
0,52 bc
|
0,49 bc
|
0,59 b
|
<0,0001
|
0,0056
|
ns
|
0,0446
|
|
K (g)
| |
12,16 a
|
4,66 b
|
5,80 b
|
4,42 b
|
3,85 b
|
6,00 a
|
<0,0001
|
<0,0001
|
0,0184
|
ns
|
|
Ca (g)
| |
1,73 a
|
0,30 c
|
0,38 c
|
0,26 c
|
0,19
c
|
1,08 b
|
<0,0001
|
<0,0001
|
0,0001
|
ns
|
|
Mg (g)
| |
0,42 a
|
0,06 d
|
0,15 c
|
0,19 bc
|
0,28 b
|
0,27 b
|
<0,0001
|
<0,0001
|
<0,0001
|
0,0071
|
|
S (g)
| |
0,41 a
|
0,13 d
|
0,20 cd
|
0,18 cd
|
0,25 bc
|
0,33 b
|
<0,0001
|
<0,0001
|
0,0004
|
ns
|
|
Fe (mg)
| |
452 a
|
48 d
|
115 c
|
155 c
|
226 b
|
52 d
|
<0,0001
|
ns
|
<0,0001
|
0,0846
|
|
Cu (mg)
| |
4,0 a
|
1,2 c
|
1,4 c
|
1,0 c
|
0,9 c
|
2,9 b
|
<0,0001
|
<0,0001
|
0,0107
|
ns
|
|
Zn (mg)
| |
14,2 a
|
2,5 b
|
2,9 b
|
2,1 b
|
1,9 b
|
14,7 a
|
<0,0001
|
<0,0001
|
0,0107
|
ns
|
|
Mn (mg)
| |
41,9 b
|
6,5 c
|
6,9 c
|
4,7 c
|
5,5 c
|
78,2 a
|
<0,0001
|
<0,0001
|
0,0051
|
ns
|
|
B (mg)
| |
5,9 a
|
1,8 c
|
3,4 b
|
3,2 b
|
5,0 a
|
3,3 b
|
<0,0001
|
0,0166
|
<0,0001
|
ns
|
|
Al (mg)
| |
22,6 e
|
155,8 cd
|
280,3 a
|
261,2 ab
|
208,4 bc
|
111,2 cd
|
<0,0001
|
ns
|
ns
|
0,0042
|
|
Valores con
una misma letra en cada fila no difieren estadísticamente según prueba de
diferencia mínima significativa al 5 %, ns = no significativo.
|
La presencia del Al redujo en más de 60% la absorción de K. El K asimilado por la planta fue similar para todas las dosis de Mg ya que produjeron la misma cantidad de biomasa. El incremento de pH de 3 a 5 mejoró significativamente la absorción del K (Marschner, 1995; Mengel & Kirkby, 2000; Villalobos & Killorn, 2001) debido a que este cambio mejoró el proceso de acumulación de biomasa. Se encontró efecto lineal negativo en la absorción de K al incrementar el contenido de Mg en solución, consecuencia del conocido antagonismo Mg/K (Lacoeuilhe & Martin-Prevel, 1971; Lahav & Turner, 1992; Turner, 1972).
La presencia de Al redujo la absorción de Ca en 83% debido a que el Al ocupa los sitios de absorción del Ca (Matsumoto, 2002). El incremento en la dosis de Mg de 25 a 50 mg l-1 mejoró levemente la absorción de Ca, pero las diferencias observadas entre las diferentes dosis de Mg no fueron significativas. Se encontró un efecto lineal negativo entre el Ca absorbido y la concentración de Mg en solución que nuevamente se puede explicar por el efecto del antagonismo Mg/Ca (Lacoeuilhe & Martin-Prevel, 1971; Lahav & Turner, 1992; Turner, 1972). El incremento de pH de 3 a 5 mejoró significativamente la absorción del Ca, probablemente porque disminuyó la concentración de Al+3 en la solución nutritiva (Marschner, 1995; Mengel & Kirkby, 2000; Villalobos & Killorn, 2001).
La absorción de Mg disminuyó en 86% con la presencia de Al en la solución, debido probablemente a la competencia por los sitios de absorción y transportadores de la membrana celular (Bose et al., 2011; Rengel & Robinson, 1989). La absorción de Mg se incrementó al aumentar la dosis de Mg en la solución, aunque no llegó a alcanzar la magnitud del Mg absorbido por las plantas del tratamiento sin Al (Tabla 5), lo anterior sugiere que existe fuerte competencia entre ambos elementos por los sitios de absorción y los transportadores de la membrana celular. Los cationes Al+3 y Mg+2 tienen similar radio de hidratación (0,48 nm para Al+3 y 0,428 nm para Mg+2) lo que favorece la competencia por los sitios de acople metal-enzima (Bose et al., 2011; Marschner, 1995; Matsumoto, 2002; Rengel & Robinson, 1989). El Al redujo la absorción de S en 68% debido a la menor producción de materia seca. La absorción de S presentó un comportamiento similar a la del Mg, debido a que la absorción mejora al incrementar la dosis Mg en solución, lo cual era de esperarse por al efecto del ión acompañante ya que la fuente de Mg utilizada fue sulfato de magnesio.
La absorción de Fe disminuyó casi en 90% al adicionar Al en la solución nutritiva, probablemente debido a la disminución en la producción de biomasa y al efecto de competencia por cargas en los sitios de absorción a nivel de raíces. Al incrementar las dosis de Mg aumentó la absorción de Fe, aunque ésta fue inferior el valor absorbido por el tratamiento testigo sin Al. La absorción de Cu, Zn y Mn tuvo un comportamiento muy similar, la aplicación de Al redujo en forma significativa la absorción de estos nutrimentos, el incremento en la dosis de Mg disminuyó aún más la absorción de estos nutrimentos, probablemente debido al efecto antagónico del Mg con estos tres cationes.
4. Conclusiones
La presencia de
Al en la solución afectó la absorción de agua y el crecimiento de la planta de
banano, situación que se reflejó en la reducción de la magnitud de las
variables de crecimiento, siendo la
acumulación de biomasa por las raíces de primer
orden la más afectada. La utilización de Al causó síntomas de toxicidad
que consistieron en la clorosis de hojas viejas y la necrosis de los bordes y
ápices de hojas, las plantas se tornaron compactas y arrepolladas, tuvieron
menor altura y acumularon menos biomasa. La dosis de 50 mg l-1 de Mg
disminuyó parcialmente el efecto
negativo de la toxicidad del Al+3 al mejorar en forma significativa la absorción de agua, la altura de planta
y la circunferencia del pseudo tallo. Dosis mayores de Mg no mejoraron la
expresión del efecto en estas variables con respecto a la dosis más baja,
debido al incremento de la conductividad eléctrica de la solución nutritiva
causado precisamente por las dosis más elevadas
de Mg y S. El incremento del pH de 3 a 5 en presencia de Al fue más efectivo
para reducir los efectos perjudiciales del Al que el aumento de las dosis de
Mg. Lo anterior debido probablemente a una menor concentración de Al+3
en solución, ya que a pH 3 este
elemento se encuentra en mayor proporción como Al+3 que es más
reactivo, mientras que a pH 5 existe
un equilibrio entre Al+3 y Al(OH)+2.
La presencia del Al en la solución nutritiva
redujo la absorción de todos los nutrimientos, el orden de reducción de los
macronutrimentos fue Mg ≥ Ca > S
> N ≥ K > P y el de los micronutrimentos fue Fe > Mn > Zn > Cu =
B. Las dosis más altas de Mg mejoraron
la absorción de P, Mg, S, el P por el
sinergismo P-Mg y el S por efecto del ión acompañante,
pero disminuyeron la absorción del Ca
y K por el antagonismo Mg/Ca y Mg/K.
Referencias
Alva, A., Edwords, D., Asher, C., & Blamey, F. (1986). Relationships between root length of soybean and calculated activities of aluminium monomers in nutrient solutions. Soil Sci. Soc. Am. J., 50, 959-962.
Ayarza, M. A., & Salinas, J. G. (1982). Estudio comparativo de la tolerancia al aluminio en tres leguminosas forrajeras. Suelos Ecuatoriales, 12(1), 110-126.
Baligar, V. C., Beaver, W. V., & Ahlrichs, J. L. (1998). Nature and distribution of acid soils in the world. Paper presented at the Workshop to development a strategy for collaborative research and dissemination of technology in sustainable crop production in acid savannas and other problem soils of the world, Indiana, USA.
Bertsch, F. (1995). La fertilidad de los suelos y su manejo. San José, Costa Rica: ACCS.
Bose, J., Babourina, O., & Rengel, Z. (2011). Role of magnesium in alleviation of aluminium toxicity in plants. Journal of Experimental Botany, 62(7), 2251-2264.
CORBANA. (2011). Estadísticas de exportación bananera 2010 (pp. 734). San José, Costa Rica: CORBANA.
Cronan, C., & Grigal, D. (1995). Use of calcium/aluminium ratios as indicators of stress in forest ecosystems. Journal of Environmental Quality, 24(2), 209-226.
De Oliveira, S. L., Coelho, E. F., & Borges, A. L. (2000). Irrigação e fertirrigação. In Z. J. Cordeiro (Ed.), Banana: produção, aspectos técnicos (pp. 60-72). Brasilia, Brasil: EMBRAPA Comunicação para Transferência de Tecnologia.
Delvaux, B. (1995). Soils. In S. Gowen (Ed.), Bananas and plantains (pp. 230-257). Londres, Inglaterra: Chapman & Hall.
Fassbender, H., & Bornemisza, E. (1987). Química de suelos con énfasis en suelos de América Latina (2da ed.). San José, Costa Rica: IICA.
Foy, C. D. (1984). Physiological effects of hydrogen, aluminium and manganese toxicities in acid soils. In F. Adams (Ed.), Soil acidity and liming (2da ed., pp. 57-97). Madison, Wisconsin: ASA.
Gaugel, C., Sierra, F., & Arevalo, G. (2003). The problem of banana root deterioration and its impact on production: Latin American experience. Paper presented at the Simposio Internacional del Sistema Radical del Banano: hacia un mejor conocimiento para su manejo productivo, San José, Costa Rica.
Gil, F. (1995). Elementos de fisiología vegetal. Madrid, España: Mundi Prensa.
Jaramillo, R., Alvarado, A., Chaverri, B., & Vázquez, A. (1979). Especificaciones teóricas para la determinación de la aptitud de las tierras para el cultivo del banano. Documento preliminar (pp. 7). San José, Costa Rica: ASBANA.
Jaramillo, R., & Vázquez, A. (1900). Manual de procedimientos para la presentación y realización de estudios detallados de suelos y clasificación de tierras para el cultivo del banano. Edición revisada del Sistema de Clasificación para determinar la aptitud de las tierras para el cultivo del banano de Jaramillo, R. & Vázquez, A. 1980 (pp. 25). San José, Costa Rica: Departamento de Investigaciones Agrícolas, CORBANA.
Kass, D. (1996). Fertilidad de suelos. San José, Costa Rica: EUNED.
Lacoeuilhe, J., & Martin-Prevel, P. (1971). Culture sur milieu artificial. Carence en K, Ca, Mg, chez le bananier : analyse foliaire. Fruits, 26(243-253).
Lahav, E., & Turner, D. (1992). Fertilización del banano para rendimientos altos (2da ed.). Berna, Suiza: International Potash Institute.
Lorenz, O., & Maynard, D. (1988). Knotts handbook for vegetable growers (3rd ed.). New York, USA: Wiley.
Marschner, H. (1995). Mineral nutrition of higher plants (2nd ed.). Londres, England: Academic Press.
Martin-prevel, P. (1980). La nutrition minerale du bananier dans le monde. Fruits, 35, 583-593.
Matsumoto, H. (2002). Plant Root Under Aluminium Stress: Toxicity and tolerance. In Y. Waisel, A. Eshel, & U. Kafkafi (Eds.), Plant roots (3rt ed., pp. 821-838). New York, USA: Hidden Half
Mengel, K., &
Kirkby, E. (2000). Principios de nutrición
vegetal (4ta ed.). Berna, Suiza: International Potash Institute.
Miyasaka, S. C.,
Hue, N. V., & Dunn, M. A. (2007).
Aluminium. In A. V. Baker & D. J. Pilbeam (Eds.), Handbook on Plant Nutrition (pp. 439-
498). Florida, USA: CRC Press.
Molina, E. (1998). Encalado
para la corrección de la acidez del suelo. San José, Costa
Rica: ACCS.
Rengel, Z., & Robinson, D. (1989).
Competitive Al+3 inhibition of net Mg+2 uptake by
intact Lolium multiflorum
roots: I Kinetics. Plant physiology, 91, 1407-1413.
Ritchey, K., Sousa, D., & Rodríguez, G. (1988). Testes biológicos
para diagnóstico da deficiencia de
calcio e toxicidade de alumínio em solos. Revista Brasilera
de Ciencia do Solo, 12, 113-120.
Robinson,
J. C. (1996). Bananas and plantains. Oxon, Inglaterra: W.K.Kellogg.
Rodríguez-García, J., Rivera, E., & Abruna, F. (1985).
Crop response to
soil acidity factors in ultisols and oxisols in Puerto Rico, 14 Plantains and Banana. Journal
of Agricultural of University of
Puerto Rico, 69, 377-382.
Rodríguez,
W., & Leihner, D. (2006). Análisis del crecimiento
vegetal. In E. Villalobos (Ed.), Fisiología
Vegetal (pp. 37). San José, Costa
Rica: Editorial de la Universidad de Costa Rica.
Rufyikiri, G., Nootens, D., Dufey, E., & Delvaux, B. (2000). Effect
of aluminium on bananas (Musa
ssp,) cultivated in acid solutions, Part I, Plant growth
and chemical composition. Fruits, 55(6), 367-379.
Rufyikiri, G., Nootens, D., Dufey, E., & Delvaux, B. (2001). Effect of aluminium on bananas (Musa ssp,) cultivated in acid solutions,
Part II, Water and Nutrient uptake. Fruits,
56(6), 5-16.
Serrano, E. (2003). Acidificación de los suelos bananeros de Costa Rica y su relación con la toxicidad de aluminio en
las raíces del cultivo de banano. Paper presented at the Simposio Internacional del Sistema Radical del Banano: hacia un mejor conocimiento para su manejo productivo,
San José, Costa Rica.
Sharrock, S., & Frison, E. (1999). Musa
production around the
world – varieties and
regional importante.
Networking banana and
plantain. In INIBAP
(Ed.), INIBAP annual report 1998
(pp. 42-47). Montpellier,
Francia: INIBAP, International
Network for the Improvement of Banana and Plantain.
Spain, J., Francis, C., Howeler, R., & Calvo, F. (1975). Differential species and varietal
tolerance to soil acidity in tropical crops and pastures.
In E. Bornemisza & A. Alvarado
(Eds.), Soil management in tropical America (pp. 308-329). Raleigh, USA: North Caroline
State University.
Stover, R. H., & Simmonds, N. W. (1987). Bananas (3rd ed.). New York,
USA: Wiley.
Tang, H., Houben, V., Nzok Mbouti, C., & Dufey, E. (1993). Diagnostic précoce de la résistance de cultivars de riz (Oriza sativa L,) á la toxicité aluminique. Agronomie, 13, 853-860.
Tisdale,
S., Nelson, W., Beaton, J., &
Havlin, J. (1993). Soil fertility and fertilizers (5th ed.). Columbus, USA: Mc Millan.
Turner, D. (1972). Banana plant growth. 1. Gross morphology. 2. Dry matter production leaf area and growth analysis. Australian Journal Experimental Agric Anim. Husb., 12(209-224).
Turner, D., & Lahav, E. (1983).
The growth of banana plants in
relation to temperature. Australian
Journal Plant Physiology, 10(1), 43-53.
Van, J., Costa de Macedo,
C., Kinet, J. M., & Bouharmont, J. (1997). Selection of Al-resistant plants from a sensitive rice
cultivar, using somaclonal variation in vitro and hydroponic cultures. Euphytica, 97, 303 -310.
Villalobos, E.,
& Killorn, R. (2001). Nutrición mineral. In E. Villalobos (Ed.), Fisiología de la producción de los cultivos tropicales (pp. 161- 196.). San José, Costa Rica: Editorial Universidad de Costa Rica.
Voight,
P. W., Godwin, H. W., & Morris, D. R. (1999). Effect of four acid soils on
root growth of clover seedlings using a soil-on-agar procedure. Plant and Soil, 205, 51-56.
von Uexküll, H., & Mutert, E. (1995). Global extent, development and economic impact of acid soils. Plant and Soil, 171, 1-15.
Westerman,
R. L. (1990). Soil testingand plant
analysis (3rd ed.). Madison, Wisconsin, USA: Soil Science
Society of America.
Wilson,
G. F., Swenenen, R., & de Langhe, E. (1987). Effects of mulch and fertilizer on yield and longevity of a medium and
giant plantain and a banana cultivar. Paper presented at the Proceedings of
the third meeting. International Cooperation for effective plantain and banana
research, Abidjan, Costa de Marfil.
Zapata, R. (2004). Química
de la acidez del suelo.
Medellín, Colombia: Universidad Nacional
de Colombia.
Zhang, X., & Jessop, R. S. (1998). Analysis of genetic
variability of aluminium tolerance response in triticale. Euphytica,
102, 177-182.




