Resumen: Justificación: El estudio de los polimorfismos de las regiones hipervariables I y II (HVI y HVII) del ADN mitocondrial (ADNmt) se ha convertido en una herramienta invaluable para la ciencia forense, ya que en algunas ocasiones un determinado individuo puede presentar más de un tipo de ADN mitocondrial este fenómeno es conocido como Heteroplasmia. Esta coexistencia de dos o más poblaciones de ADNmt puede ocurrir en una sola mitocondria, célula o individuo, lo que puede aumentar la complejidad en la interpretación de los resultados de las experticias forenses. Objetivos: Analizar la frecuencia de la heteroplasmia en las regiones HVI y HVII del genoma mitocondrial en una muestra de la población de Maracaibo. Metodología: Se seleccionaron al azar 50 muestras de ADN de la población de Maracaibo, las regiones hipervariables se amplificaron mediante reacción en cadena de la polimerasa, posteriormente se secuenciaron mediante método de Sanger y los fragmentos se separaron por electroforesis capilar, se reportaron las diferencias con respecto a la secuencia de referencia de Cambridge. Resultados: El 26% de las muestras presentaron heteroplasmia en la región HVI, el 52% en la región HVII. Conclusiones: El hecho de aparecer la heteroplasmia en una determinada secuencia no inválida el uso del análisis del ADN mitocondrial con fines forenses, dependiendo de la complejidad del caso a peritar la heteroplasmia puede ser de gran ayuda.
Palabras clave: Regiones hipervariables,Heteroplasmia, ADN mitocondrial.
Abstract: Background: The study of the polymorphisms of the hypervariable regions I and II (HVI and HVII) of the mitochondrial DNA (DNAmt) has become an invaluable tool for forensic science, in some occasions a certain individual can present more than one type of mtDNA, this phenomenon is known as Heteroplasmy, this coexistence of two or more populations of DNAmt can occur in a single mitochondrion, cell or individual, which can increase the complexity in interpreting the results of forensic expertise. Objectives: Analyze the frequency of heteroplasmy in the HVI and HVII regions of the mitochondrial genome of 50 randomly selected DNA samples from the Maracaibo population. Methodology: The hypervariable regions were amplified by PCR, then sequenced using the Sanger method and the fragments were separated by capillary electrophoresis, differences were reported with respect to the Cambridge reference sequence. Results: 26% of the samples presented heteroplasmy in the HVI region, while in the HVII region it was 52%. Conclusions: The appearance of heteroplasmy in a certain sequence does not invalidate the use of mitochondrial DNA analysis for forensic purposes, depending on the complexity of the case to assess heteroplasmy can be of great help.
Keywords: Hypervariable regions, Heteroplasmy, Mitochondrial DNA.
Artículos Originales
Frecuencia de heteroplasmia en las regiones hipervariables HVI y HVII del ADN Mitocondrial en una muestra de la población de Maracaibo, Venezuela
Recepción: 05 Julio 2019
Aprobación: 19 Julio 2019
Autor de correspondencia: jmquintero@fmed.luz.edu.ve

Esta obra está bajo una Licencia Creative Commons Atribución-NoComercial 4.0 Internacional.
El genoma mitocondrial está conformado por una doble cadena de ácido desoxirribonucleico (ADN) circular de 16569 pares de bases (pb) y organizado en dos regiones clasificadas según su función en una región codificante que contiene 37 genes implicados principalmente en la fosforilación oxidativa y una región no codificante exenta de genes denominada región control o asa de desdoblamiento1,2.
La región control (RC) consta de 1121 pares de bases de longitud, abarca desde la posición 16024 hasta la 16569 continuando desde la posición 1 hasta la 5763. Es precisamente esta región la que es ampliamente analizada por los laboratorios de genética forense y antropología molecular a nivel mundial debido a que no está involucrada en la codificación de un producto génico 4,5, permitiendo que se generen mutaciones en ella, acumulándose con una frecuencia de 5 a 10 veces mayor que el resto del ADNmt 6, en esta región se encuentra la mayor variabilidad interpersonal ya que la probabilidad de que dos individuos no relacionados de una población posean la misma secuencia en la RC es menor al 1 % (dependiendo de la población) 7.
Estas mutaciones se concentran en tres regiones específicas que según su grado de variabilidad son: la región hipervariable 1 (HVI) que abarca las posiciones 16024 a la 16365, la región hipervariable 2 (HVII) que va desde la posición 73 a la 340 y la región hipervariable 3 (HVIII) que va de la posición 440 a la 560, además existen otras regiones con un menor grado de variabilidad conocidas como región variable 1 (VRI) localizadas desde la posición 16366 hasta la 72 y región variable 2 (VRII) que está comprendida entre las posiciones 341 y 576.
El tipo de mutación más común en estas regiones es la puntual, con una frecuencia de 40 veces de tipo transición (sustitución de una base púrica por otra púrica o pirimidínica por pirimidínica) a una de tipo transversión (cambio de una base nucleotídica púrica por pirimidínica o viceversa) y pequeñas inserciones y deleciones en regiones homopolimericas (poliC) presentes en ambas regiones hipervariables, entre las posiciones 303 y 315 y entre 16183 y 16194 8,9.
Ahora bien, el ADNmt no solo posee un elevado grado polimórfico en la región control, ostenta además una serie de características propias que hacen de él una estupenda herramienta en casos forenses complejos, donde a través del análisis del ADN nuclear no sería posible obtener algún resultado concluyente. Entre las características más importantes están: Herencia materna, alto número de copias, alta tasa de mutación y la heteroplasmia.
En algunas ocasiones, un determinado individuo, por efecto de las mutaciones, puede presentar más de un tipo de ADNmt, este fenómeno es conocido como heteroplasmia (para diferenciarlo del estado normal de homoplasmia), esta coexistencia de dos o más poblaciones de ADNmt puede ocurrir en una sola mitocondria, célula o individuo 9,10.
Durante el desarrollo las moléculas de ADNmt se replican independientemente unas de otras y este proceso no está estrictamente ligado a los procesos de división celular meiótica y mitótica. Por otra parte, el proceso de replicación del ADNmt está asociado a un mayor nivel de error que el que se encuentra en el ADN nuclear, se piensa que los mecanismos de reparación actúan deficientemente en las mitocondrias La teoría de cuello de botella explicaría en cierta manera el porqué de la aparición de diferentes proporciones de heteroplasmias en distintos tejidos de un mismo individuo, así como la aparición de estas diferencias entre individuos relacionados por vía materna11. Cuando aparece heteroplasmia, lo habitual es que los tipos de ADNmt que aparecen en un individuo difieran en una sola base, siendo infrecuente la heteroplasmia en dos o más sitios. Se ha comprobado que tejidos como los pelos son más susceptibles a que aparezca el fenómeno de la heteroplasmia 12 y que es más frecuente en la población de lo que se pensaba 13. La aparición de heteroplasmias ha sido recogida en diversos casos de interés forense, siendo quizás el más conocido el caso de identificación de los restos óseos del Zar Nicolás II 14.
La heteroplasmia se puede dividir en heteroplasmia de secuencia que ocurre cuando en una posición determinada de la secuencia del ADNmt hay más de una base, por ejemplo la existencia de C/T en una determinada posición del ADNmt y heteroplasmia de longitud que se manifiesta como la variación en el número de bases existentes en un fragmento homopolímerico, por ejemplo es característica la variación en el número de citosinas en las regiones localizadas entre las bases 16183 y 16194 en HVI y entre las bases 303 y 315 en HVII, es común observar este tipo de heteroplasmia como un corrimiento del marco de lectura en el electroferograma. Así, en un mismo individuo, pueden coexistir poblaciones de ADNmt las cuales difieren en el número de citosinas dentro de esos fragmentos 9.
Cuando una célula heteroplásmica se divide, la transmisión de las mitocondrias a las células hijas se realiza al azar, resultando que la proporción de ADNmt mutante y normal, tras varios ciclos de división, puede derivar hacia el mutante o el normal en un proceso conocido como segregación replicativa. Esto hace que los niveles de heteroplasmia no siempre sean los mismos en los diferentes tejidos de un individuo 15.
El hecho de aparecer la heteroplasmia en una determinada secuencia no inválida el uso del análisis del ADN mitocondrial con fines forenses, sin embargo, se debe verificar que efectivamente se trata de una heteroplasmia. Para ello se debe confirmar la aparición de dicho evento tanto en la cadena 5`-3` como en la correspondiente cadena complementaria 3`-5` de la secuencia analizada. En este caso la lectura de los picos en el electroferograma debe ser clara y por encima del nivel basal o de ruido, concretamente un 20% por encima de ese nivel basal, sin dejar lugar a dudas de que efectivamente se trata de una heteroplasmia. Si no existe dicha certeza conviene confirmarlo con adicionales análisis. Cuando aparece una heteroplasmia puede ocurrir que las poblaciones de mitocondrias que coexisten lo hagan en relación 80:20%, en cuyo caso los picos correspondientes a las bases que generan la heteroplasmia, definirán una posición donde pudiera ser difícil determinar con exactitud de que base se trata. En otras ocasiones dichas poblaciones coexisten en porcentajes variados, esta diferencia se suele reflejar también en el electroferograma, dependiendo de la complejidad del caso a peritar la heteroplasmia puede ser de gran ayuda 14.
Población y muestra: La muestra comprendió 50 individuos, sin relación de parentesco, residentes en la ciudad de Maracaibo, estado Zulia, Venezuela; seleccionados independientemente de la edad y sexo de los individuos. Todas las personas autorizaron el uso de su perfil genético para estudios genético-poblacionales, una vez informado del compromiso institucional de mantener los datos anónimos.
Extracción de ADN, amplificación mediante PCR y caracterización por electroforesis capilar: El ADN de cada muestra sanguínea se extrajo previamente mediante la técnica Salting Out modificada 16, se amplificaron a través de la PCR (del inglés, Polymerase Chain Reaction) y por separado las regiones HVI (nucleótidos 16024 a 16365) y HVII (nucleótidos 72 a 340) del ADNmt. La mezcla de la PCR estuvo constituida por: 10 μl de buffer Taq polimerasa 10X; 1,5 μl MgCl2 (1,5 mM); 1 μl dNTPs (0,2 mM); 0,4 μl Taq polimerasa (0,5 unidades); 28,1 μl de Agua; 2 μl (15 pmol) de cada iniciador 17, en conjunto con 1 ng de ADN genómico en un volumen final de 50 μl. Las condiciones de amplificación de la PCR fueron: 95°C 11 min; [94°C 45 s; 50°C (iniciadores HVI) 55ºC (iniciadores HVII) 1 min; 72°C 5 min 30 s; 30 veces]; 72°C 45 s; 4°C.
Los productos amplificados fueron purificados mediante el estuche comercial Wizard® PCR Preps DNA Purification System (Promega® Corporation, Madison, WI, USA), luego fueron secuenciados mediante BigDye v3.1 Ready Reaction (ThermoFisher Scientific, Waltham, MA, USA), las muestras secuenciadas fueron purificadas utilizando Wizard® MagneSil® Sequencing Reaction Clean-Up System (Promega® Corporation, Madison, WI, USA). Las muestras ya purificadas fueron caracterizadas en el equipo semiautomatizado ABI PRISM® 310 Genetic Analyzer (ThermoFisher Scientific, Waltham, MA, USA), el cual se acondicionó previamente utilizando POP-6 (ThermoFisher Scientific, Waltham, MA, USA) y capilar de secuenciación de 61 cm de longitud por 50 μm de diámetro, bajo los siguientes parámetros de corrida: Tiempo de inyección: 50 s, temperatura 50ºC y tiempo de corrida para HVI de 90 min y HVII 70 min. Los datos crudos de la secuenciación se analizaron con el programa Sequencing Analysis (ThermoFisher Scientific, Waltham, MA, USA), luego, se procedió a la correspondiente edición con el programa Sequence Scanner v1.0 (ThermoFisher Scientific, Waltham, MA, USA). Finalmente se procedió a la comparación de los fragmentos secuenciados con la secuencia nucleotídica de referencia rCRS (GenBank NC_012920.1), utilizando el software Staden Package v2.0.0b9 18, los haplogrupos fueron asignados mediante el software on-line mtDNA manager 19.
La clasificación de la heteroplasmia se realizó mediante el método de Lutz-Bonengel y colaboradores20 aplicando las siguientes fórmulas:
rseq = rQV (16189-16207.x) / rQV (16170-16188.x)*
rseq = rQV (310-328.x) / rQV (291-309.x)
Siendo rseq la relación de los valores de calidad y rQV los valores de calidad de cada nucleótido.
*Fórmula adaptada a la región HVI usada para esta investigación, no aportada por Lutz-Bonengel.
Los resultados de las fórmulas permiten clasificar las secuencias en cuatro grupos: Grupo I (si la relación es >1, homoplasmia), Grupo II (0,85 - 0,99), Grupo III (<0,85), y Grupo IV (patrón desordenado de picos), como se muestran en la Figura 1.
La presente investigación se realizó siguiendo los lineamientos de la Sociedad Internacional de Genética Forense para la caracterización del ADNmt 21.
Figura 1. Clasificación mediante secuenciación directa de las heteroplasmias de longitud.
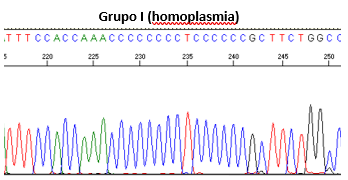
Figura 1: Clasificación mediante secuenciación directa de las heteroplasmias de longitud.
Grupo I (homoplasmia)
Autor
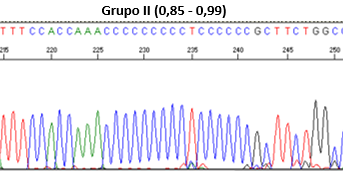
Figura 1. Clasificación mediante secuenciación directa de las heteroplasmias de longitud.
Grupo II (0,85 - 0,99)
Autor
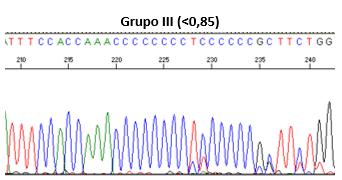
Figura 1. Clasificación mediante secuenciación directa de las heteroplasmias de longitud.
Grupo III (<0,85)
Autor
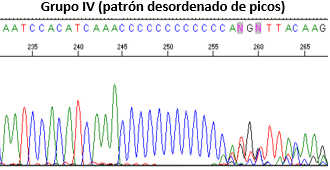
Figura 1. Clasificación mediante secuenciación directa de las heteroplasmias de longitud.
Grupo IV (patrón desordenado de picos)
Autor
En 50 muestras de ADN analizadas, considerando los segmentos poliC (entre las posiciones 16183 y 16194 en la región HVI y entre 303 y 315 en HVII), para la región HVI, se detectaron 65 sitios variables y 37 haplotipos diferentes, de las cuales 32 fueron únicos (frecuencia haplotípica 0,02 para cada uno) y el resto se encuentra en al menos dos muestras. Mientras que para la región HVII, 43 posiciones nucleotídicas variables fueron detectadas y 28 haplotipos diferentes fueron observados, de los cuales 22 fueron únicos (frecuencia haplotípica de 0,02 en cada uno), como se muestra en el Cuadro I. En combinación ambas regiones hipervariables presentaron 108 polimorfismos y 43 haplotipos distintos, de los cuales 38 fueron únicos 4.
Haplotipos mitocondriales de las regiones hipervariables I y II de individuos de la ciudad de Maracaibo, Venezuela.

M: Muestra; Hg: Haplogrupo, cuadro construido por autores.
En las muestras solo se observó heteroplasmia de longitud, esta se clasifico mediante secuenciación directa en cuatro grupos I, II, III y VI según el grado en la cual se presentó (Figura 1). El 26% de las muestras presentaron heteroplasmia en la región HVI, mientras que en la región HVII fue mayor con 52% del total se secuencias analizadas. Treinta y un (31) muestras del total presentaron heteroplasmia en al menos una región hipervariable, por lo que la heteroplasmia estuvo presente en el 62% de las muestras, (Cuadro II).
Clasificación de la heteroplasmia de longitud en las regiones HVI y HVII.

Autores
Una transición en particular la 16189T>C origina un fragmento poliC de 10 citosinas entre los nucleótidos 16183 y 16194, al ocurrir esto generalmente la ADN polimerasa mitocondrial gamma comete errores de tipo “slippage”, habitualmente insertando un número variable de citosinas en esta región, esto ocasiona una superposición de estos fragmentos, originando que en los electroferogramas en este segmento se observe como un “ruido de fondo” 22, dificultando la lectura e interpretación de las secuencias en esa región, el 26% de las muestras analizadas mostraron este cambio de T por C en la región HVI, este porcentaje varía en las diferentes poblaciones encontrando 5% en Suiza 23, 15% Austria 24 y 37% en Japón 25.
Un estudio realizado por Irwin y colaboradores donde se secuenció la región control de 5,015 individuos de diversas poblaciones, arrojó que el 52% de esas muestras presentaban heteroplasmia de longitud, y esto era observado claramente cuando se producía un segmento poliC generalmente de ≥9 citosinas, que se originaba al ocurrir las transiciones en las posiciones 16184 y 16198 en la región HVI, causando una mezcla en el 97% de los casos de 10 y 11 citosinas, la región HVII en cambio, mostró que el número de citosinas necesario para provocar el “slippage” era menor que en HVI de ≥7 citosinas y el 76% de los casos mostró una mezcla de 8 y 9 citosinas 26. Ese mismo comportamiento ocurrió en las muestras de Maracaibo, las muestras que presentaron los cambios nucleotídicos 16183A y 16189C (ejemplo la muestra MBO0284) produjeron un fragmento de aproximadamente 12 citosinas, presentaron heteroplasmia grado IV, mientras que la región HVII no hubo cambios favorecieran la formación de ≥7 citosinas por lo que ninguna de las secuencias presentó una heteroplasmia grado IV.
Por otro lado, 12 de las 16 muestras mencionadas anteriormente pertenecen al grupo IV, el cual se asocia a segmentos poliC de 11, 12, 13 y 14 citosinas en una misma muestra, todas mostraron la transversión 16183A>C, la transición 16189T>C y la inserción 16193.1C, este grupo se caracteriza técnicamente por una caída de la lectura electroforética después del fragmento poliC, haciendo prácticamente ilegible la secuencia después de ese fragmento por lo que la única manera de leerla es a través del fragmento complementario forward o reverse según el caso.
Un segmento parecido al de la región HVI está presente en la HVII entre los nucleótidos 303 y 315, pero a diferencia la región HVI, en la región HVII generalmente no sucede el cambio T por C en este caso en el nucleótido 310, y efectivamente ninguna de las muestras del presente trabajo presenta ese cambio, sin embargo, la heteroplasmia estuvo presente en un porcentaje mayor que en HVI en un 52% de las muestras, exceptuando las que clasificaron dentro del grupo I, la proporción de muestras fue igual para los grupos II y III,14 muestras en cada grupo y ninguna muestra se clasificó dentro del grupo IV. El grupo II se asocia a segmentos poliC de 7 y 8 citosinas mientras que el grupo III a 9 y 10 citosinas 20.
Los grupos II y III son grupos que presentan menos complejidad que el grupo IV, debido a que no hay un “ruido de fondo” tan marcado como el grupo IV, no obstante, se corroboro la presencia de la heteroplasmia en dos secuenciaciones independientes al igual que en la región HVI tal como lo establece la Sociedad Internacional de Genética Forense 9,24.
Ahora bien, es importante destacar que la presencia de heteroplasmia puede ser de gran ayuda como en el caso del Zar Nicolás II, donde Georgij Romanov el hermano del Zar presentaba heteroplasmia de posición en el nucleótido 16169 al igual que el Zar Nicolás II; sin embargo, dada la complejidad de la interpretación de la heteroplasmia, han surgido recomendaciones para la asignación coherente de las secuencias con heteroplasmia de longitud, las cuales fueron seguidas en esta investigación 27, debido a esta complejidad algunos trabajos omiten esta región en sus resultados.
Conclusiones
Es importante determinar la frecuencia de la heteroplasmia en las diversas poblaciones ya que puede acrecentar la complejidad en la interpretación de los resultados en las experticias forenses, por lo que deben tomarse estrategias (cambiar iniciadores, uso de plásmidos, entre otros) que permitan obtener resultados óptimos para ser valorados por el genetista forense, por eso motivo las regiones poliC son generalmente obviadas en los reportes, a pesar de ello, es indiscutible la utilidad del genoma mitocondrial en casos forenses donde a través de otras estrategias moleculares no sería posible obtener resultados concluyentes.
CITAR COMO: Quintero JM, Pardo T, Borjas L. Frecuencia de heteroplasmia en las regiones hipervariables HVI y HVII del
ADN Mitocondrial en una muestra de la población de Maracaibo. Rev. cienc. forenses Honduras. 2019; 5(2):14-24.
ASPECTOS
ÉTICOS: Los autores declaran que no existe conflicto de interés en
la publicación de este artículo.
Todos los participantes firmaron consentimiento informado
autorizando la participación en el estudio.
SLAGF: El Dr. José Miguel quintero es el delegado por Venezuela de la Sociedad Latinoamericana de Genética Forense
http://www.bvs.hn/RCFH/pdf/2019/pdf/RCFH5-2-2019-4.pdf (pdf)
https://www.lamjol.info/index.php/RCFH/article/view/8885 (pdf)
jmquintero@fmed.luz.edu.ve
Figura 1: Clasificación mediante secuenciación directa de las heteroplasmias de longitud.
Grupo I (homoplasmia)
Autor
Figura 1. Clasificación mediante secuenciación directa de las heteroplasmias de longitud.
Grupo II (0,85 - 0,99)
Autor
Figura 1. Clasificación mediante secuenciación directa de las heteroplasmias de longitud.
Grupo III (<0,85)
Autor
Figura 1. Clasificación mediante secuenciación directa de las heteroplasmias de longitud.
Grupo IV (patrón desordenado de picos)
Autor
Haplotipos mitocondriales de las regiones hipervariables I y II de individuos de la ciudad de Maracaibo, Venezuela.
M: Muestra; Hg: Haplogrupo, cuadro construido por autores.
Clasificación de la heteroplasmia de longitud en las regiones HVI y HVII.
Autores


