Introducción
Los pastizales serranos conforman un mosaico de comunidades a nivel regional, resultante de un gradiente complejo de altitud y posición topográfica sometido a procesos geomorfológicos, edáficos, hidrológicos y de disturbios (González et al., 1999). Dicha heterogeneidad florística a escala local se relaciona con potenciales productivos diferentes (Cantero et al., 2001) lo que provoca respuestas diferenciales frente a los disturbios generados por el manejo a través del pastoreo, fuego prescripto y reemplazos por genotipos forestales exóticos (Nuñez et al., 2002; Cingolani et al., 2015).
Díaz et al., (2006) plantea que los pastizales de montaña de las sierras del centro de Córdoba tuvieron una ocupación continua por grandes herbívoros al menos desde el pleistoceno (hace aproximadamente 10.000 años), incluyendo la presencia de ungulados domésticos y semi domésticos desde al menos aproximadamente 300 años. Demostrando así que estos pastizales se caracterizan por poseer una larga historia de pastoreo.
En la actualidad el ganado bovino pastorea en lotes de gran extensión (más de 200 ha) donde coexisten diferentes comunidades de pastizales y lo hace en forma continua y liviana (0.2- 0.3 EV/ha) a lo largo de todo el año, lo que conduce a una sobreutilización de la vegetación en algunas áreas y subutilización en otras, provocando importantes desajustes tales como la generación de parches denudados y graves pérdidas de suelo por erosión hídrica en masa (Cantero et al., 1999).
El entendimiento de las relaciones entre los cambios de la vegetación y el pastoreo es fundamental para generar herramientas de manejo que puedan predecir el impacto del pastoreo sobre el mantenimiento de la diversidad y la riqueza florística de los pastizales naturales en el tiempo (Loydi et al., 2019).
En estudios más recientes llevados a cabo en los pastizales de montaña de Córdoba, Argentina, se observó que la biodiversidad se maximiza con una presión de herbivoría heterogénea dentro de cada uno de los diferentes ambientes que componen el paisaje (Cingolani et al.,2008).
En su análisis sobre las comunidades de pastizales y su resiliencia, Bakker et al., (1996) y Bastos et al., (2018), sostienen que el estudio del banco de semillas del suelo (BSS), juega un rol significante en la composición florística de las futuras comunidades vegetales, en especial, luego de un disturbio, pudiendo afectar, por un lado, la colonización de sitios perturbados y por el otro las tendencias sucesionales las cuales difieren de la vegetación existente y se reflejan por encima y debajo del suelo (Funes et al., 2001; Hudson et al., 2015).
Al BSS lo definen, Simpson et al., (1989) como el conjunto de propágulos de semillas viables en el suelo y sobre su superficie, las que pueden persistir por períodos de tiempo largos o cortos. A su vez, Bakker et al., (1991), Fenner y Thompson (2005) han coincidido en la definición de tres tipos BSS, de acuerdo a la longevidad de las semillas que lo integran y han sugerido protocolos específicos para su estudio: Banco transitorio, Banco persistente a corto plazo, Banco persistente a largo plazo. En cuanto a las metodologías de análisis del BSS Thompson et al., (1997) plantean tres métodos: de separación, de germinación y de enterramiento. En este estudio, se focaliza en el método de germinación, que se basa en la emergencia de plántulas bajo condiciones de campo, el cual registra solo las especies y los individuos que emergen en las condiciones micro-meteorológicas imperantes de una delgada capa del suelo en la cual la luz puede penetrar, ello permite ampliar el área de muestro, tratando de incluir a las especies raras y a las que tienen una distribución irregular. Estos mismos autores plantean que las especies emergidas en el campo estuvieron asociadas al banco de semillas transitorio y no a las semillas que se encontraban en las capas más profundas del suelo, clasificadas como persistentes.
Hasta la actualidad, en Argentina, no existen antecedentes sobre la realización de experimentos de este tipo, en pastizales naturales, si se ha estudiado que la composición de las comunidades reunidas de forma natural está determinada por los filtros ambientales, el reclutamiento y la dispersión de especies entre otros procesos (Díaz et al., 2003; Marberg, 2013). Experimentos similares fueron realizados en otros países, tal lo reportado por Onipchenko et al., (2004) realizados en comunidades alpinas del noroeste del Cáucaso, confirmaron que la germinación en el campo depende de condiciones microambientales asociadas al sitio y al grado de disturbio.
La composición cualitativa y cuantitativa del BSS está influenciada por diversos factores. El pastoreo es uno de ellos y actúa en la regulación de la estructura y el funcionamiento de las comunidades de pastizales naturales, y por consiguiente provoca cambios en la estructura y composición de los bancos de semillas, alterando su diversidad y densidad (Rauber et al., 2023). El pastoreo o su exclusión, influyen sobre la acumulación o la reducción de la broza existente en los pastizales, (Morici et al., 2009) y afecta la coexistencia de las especies por cambios en las jerarquías competitivas o posibilitando el reemplazo de especies (Kahmen et al., 2002, Sandoval–Calderón et al., 2024).
Las especies que incrementan su abundancia bajo la presión de la herbivoría poseen atributos que las diferencian de las que decrecen con ella (Díaz et al., 2007 y Cingolani et al., 2008). En los pastizales con pastos bajos, desarrollados en laderas y sobre suelos someros las especies tienden a arraigar en grietas y fisuras existentes en las rocas o se concentran en acúmulos edáficos (Cantero et al., 2001), la cobertura es variable, aunque nunca llega al 100%, suelen acumular broza a una tasa más lenta que los pastizales altos, debido a que generalmente son de baja estatura y de menor productividad ya que están sujetos a estrés hídrico. En este mismo sentido, en sitios pastoreados donde el suelo carece de una cubierta de mantillo, Márquez et al., (2002) observaron en el banco de semillas del suelo un aumento de las gramíneas anuales.
En cambio, el pastizal alto ocupa sustratos de profundidad variable con suelos generalmente bien drenados y se desarrollan en laderas con gradientes, suaves. En ésta comunidad, dominan las formas de crecimiento de gramíneas altas, las cuales invierten más recursos en la producción aérea que subterránea. En este pastizal, si las condiciones cambian, por ejemplo, por sobrepastoreo y fuego recurrentes, puede cambiar su fisonomía (Cingolani et al., 2015). El pastoreo liviano genera en los mismos, la acumulación de material muerto en pié o depositado sobre el suelo, el cual puede ser un factor de control, ya sea evitando la emergencia de plántulas por retención de semillas que inhibe el enterramiento de las mismas o por el macollamiento de las especies dominantes (Baskin y Baskin 2014).
De lo anterior se desprende que la remoción de la parte aérea, modifica el microclima, principalmente el espectro lumínico, altera las condiciones de crecimiento y permite la germinación de especies en los espacios entre matas (Nuñez et al., 2001). La mayor inversión en parte aérea y los múltiples estratos presentes en este pastizal sugiere que en esta comunidad con un pastoreo moderado el tamaño del BSS sería mayor al de los pastos bajos.
Con el sistema de pastoreo continuo, utilizado corrientemente en los pastizales mencionados en los párrafos anteriores, el ganado bovino pastorea con más frecuencia el pastizal bajo que el pastizal alto, dado que las especies codominantes y sus acompañantes, ofrecen una mayor calidad de forraje y un canopeo denso y bajo que resiste bien el pastoreo y rebrota rápidamente, lo que provee al ganado de forraje renovado continuamente en época de primavera-verano coincidiendo con lo planteado por Diaz et al., 2007 y Cingolani et al. 2015.
En ese sentido, Amuchástegui, (2016) plantean que en sitios pastoreados con altas cargas, la gran densidad de semillas de las dicotiledóneas presentes en el banco está relacionada con la abundancia de éstas especies no palatables de hábito rastrero y arrosetado presentes en la vegetación establecida, que se ven favorecidas por el pastoreo selectivo, resultados similares fueron observados por Márquez et al., (2002) y Funes et al., (2001) en pastizales súb-húmedos del centro de Argentina; Kahmen y Poschlod, (2008), en pastizales cálcicos europeos. Es así que las especies de forma de crecimiento en roseta, con tamaño grande de semillas, rápida germinación y desarrollo radicular, representarían una estrategia que competitivamente podría desplazar a las plántulas que no posean estos atributos (Haretche y Rodríguez, 2006).
El efecto del pastoreo prolongado sobre el banco ha sido bien estudiado (Milchunas y Laurenroth, 1993; Vecchio et al., 2022), quienes afirman que, la onda máxima de emergencia de plántulas, suele coincidir con el período de máxima frecuencia de ocurrencia de suelo desnudo (Westoby y Wright, 2006), sostienen que los parches de vegetación juegan un rol importante en la modificación de los procesos ecológicos en los pastizales. No existe un acuerdo generalizado sobre el efecto del pastoreo sobre la densidad del banco de semillas. Por un lado, la ausencia de cambios significativos en la densidad del banco ha sido observada por Milberg (1995) y por Márquez et al., (2002); en cambio, Ortega et al., (1997) y Tessema et al., (2016), hallaron un menor número de semillas en los bancos de sitios pastoreados, mientras que Marco y Páez (2000) y Minervini Silva (2020), reportaron un mayor número de semillas en BSS en sitios pastoreados.
Se ha observado que la exclusión al pastoreo, incrementa el tamaño del banco, debido a la reducción de la predación de flores y frutos, especialmente en las especies perennes (Vecchio et. al., 2019). Estos efectos pueden producir cambios en la estructura de la comunidad, en particular en su composición, riqueza y diversidad (Facelli y Facelli, 1993; Rauber et al.,2023). El pastoreo liviano o su ausencia, produce un incremento de gramíneas perennes cespitosas (Díaz et al., 2006), las que reemplazan a las especies pequeñas adaptadas a la herbivoría, permitiendo la acumulación de mantillo, el cual altera la luz incidente, la temperatura, la dinámica del agua y nutrientes del suelo, factores que afectan la germinación (Baskin y Baskin 2014). Además, Márquez et al., (2002) observaron que en sitios excluidos al pastoreo el 43% de las semillas quedaron retenidas en el mantillo, dificultando su germinación y establecimiento en el campo.
En cuanto a la presencia de especies exóticas, la baja cantidad de las mismas observadas con intensidades moderadas a altas de pastoreo (Márquez et al., 2002) en el banco de semillas, sugieren que el pastoreo no estaría actuando como una perturbación que favorecería a estas especies. Por otro lado, Amuchástegui (2016), no observó un aumento de las especies exóticas en el banco de semillas del suelo, pero sí registró un incremento en la densidad de las especies exóticas en sitios altamente pastoreados, lo cual podría indicar un deterioro del pastizal a corto plazo en la vegetación establecida, inmediatamente luego de un disturbio (Vecchio et al., 2022).
Del conocimiento generado a través de los trabajos mencionados, surgieron muchos interrogantes sobre el manejo y conservación de los pastizales serranos del sur de la provincia de Córdoba, uno de ellos fue la necesidad de disponer de un estudio de banco de semillas en condiciones de campo, a los fines de generar información que contribuya a realizar un uso más sustentable del recurso pastizal natural. Objetivo. Evaluar los efectos de la intensidad del pastoreo, sobre el tamaño y la variación de la composición florística del banco transitorio de semillas del suelo en dos comunidades de pastizales serranos centro argentinos.
Materiales y métodos
Descripción del área de Estudio
El área de estudio pertenece a la “Cuenca la Tapa” está situada en la Sierra de Comechingones, (32° 46' de latitud Sur y 64° 56' de longitud oeste). Comprende siete unidades del paisaje: planicies (altas y bajas), laderas (escarpadas, moderadas, suaves), valles y mallines (González et al., 1999). Estas unidades han sido diferenciadas por topografía (altitud y pendiente), suelo y vegetación. Dentro de esta área de estudio se escogieron para su estudio las unidades que se encuentran a mayor altitud (1400 msnm a 1800 msnm), y corresponden a planicies altas y laderas moderadamente escarpadas. El complejo de suelos denominado ¨pampas de altura¨, se encuentra ubicado por encima de los 1500 msnm. Son suelos someros de textura franca gravillosa, apoyados sobre roca alterada o roca firme. El clima del área es de tipo templado semiárido-subhúmedo con estación invernal seca, de escasa deficiencia hídrica anual y alta variabilidad en las precipitaciones, la precipitación anual promedio fue de 948mm.
Desde el punto de vista fitogeográfico, la región pertenece al Distrito Chaqueño Serrano (Cabrera 1976; Luti et al., 1979), sin embargo, Funes y Cabido (1995); Cabido et al., (1998) han sugerido su reubicación (distrito subandino) debido a la gran proporción de especies de linaje andino-patagónico presentes en el área. Desde el punto de vista florístico es la región con mayor riqueza florística del centro de Argentina, predominando las especies nativas y entre ellas muchas endémicas (Cantero y Bianco, 1986; Zuloaga et al., 1999). Según Cantero et al., (2001), los pastizales estudiados en el área exhiben una alta riqueza (368 taxones registrados).
Las diferentes comunidades están asociadas a distintas condiciones ambientales de las cuales la profundidad de suelo es el filtro ambiental más importante. Los paisajes más productivos (pampas) se encuentran por encima de los 1600 msnm (González, 2000), y presentan un predominio de pastizales de gramíneas cespitosas altas pertenecientes a los ecogrupos I y IV (Festuca hieronymi . Deyeuxia hieronymi), en combinación con una vegetación baja ecogrupo V (Sorghastrum pellitum) (Cantero et al., 1999). El área de estudio involucró principalmente a los Ecogrupos I y V. El primero de ellos se corresponde a planicies altas relativamente planas, con pendientes simples y largas de bajo gradiente (4 - 7%). La vegetación predominante pertenece a pajonales y pastizales. Se caracterizan por la dominancia de Deyeuxia hieronymi y Nassella pseudopampagrandensis, gramíneas altas, cespitosas, que dejan escasos espacios internos para el crecimiento de otras especies. El Ecogrupo V corresponde a laderas moderamente escarpadas. La vegetación predominante está dominada por Sorghastrum pellitum y Jarava juncoides, asociada a suelos poco profundos, ubicados en laderas con afloramientos rocosos y material disperso en la superficie.
Diseño Experimental
Se aplicó un diseño experimental completamente aleatorizado, el cual involucraba tres tratamientos de intensidades de pastoreo: 1 EV/ha, 0,25 EV/ha y exclusión al pastoreo doméstico (Clausura). Se siguió la definición de EV propuesta por a Coop (1965), es el promedio anual de los requerimientos de una vaca de 400 kg de peso, que gesta y cría un ternero hasta el destete a los 6 meses de edad con 160 kg de peso, incluido el forraje consumido por el ternero. Equivale a los requerimientos de un novillo de 410 kg de peso que aumenta 500 g diarios.
Al momento de iniciado este trabajo, el ganado había sido retirado con anterioridad (1 año). El estudio se llevó a cabo en las dos comunidades (sitio): Deyeuxia hieronymi (PGA): pastizal de gramíneas altas, y Sorghastrum pellitum: pastizal de gramíneas bajas (PGB) que coexisten espacialmente en cada parcela (Figura 1).
Para evaluar la emergencia de plántulas a campo (Figura 1) se ubicaron en cada tratamiento, 20 parcelas de lectura permanente de un área de 0,04m. cada una, donde se censaron y extrajeron simultáneamente las plántulas que iban apareciendo, con una frecuencia de dos (2) censos por mes durante las estaciones de otoño y primavera. Los resultados se expresaron en valores medios para un área de 0,8m., que es el área muestreada en cada tratamiento.
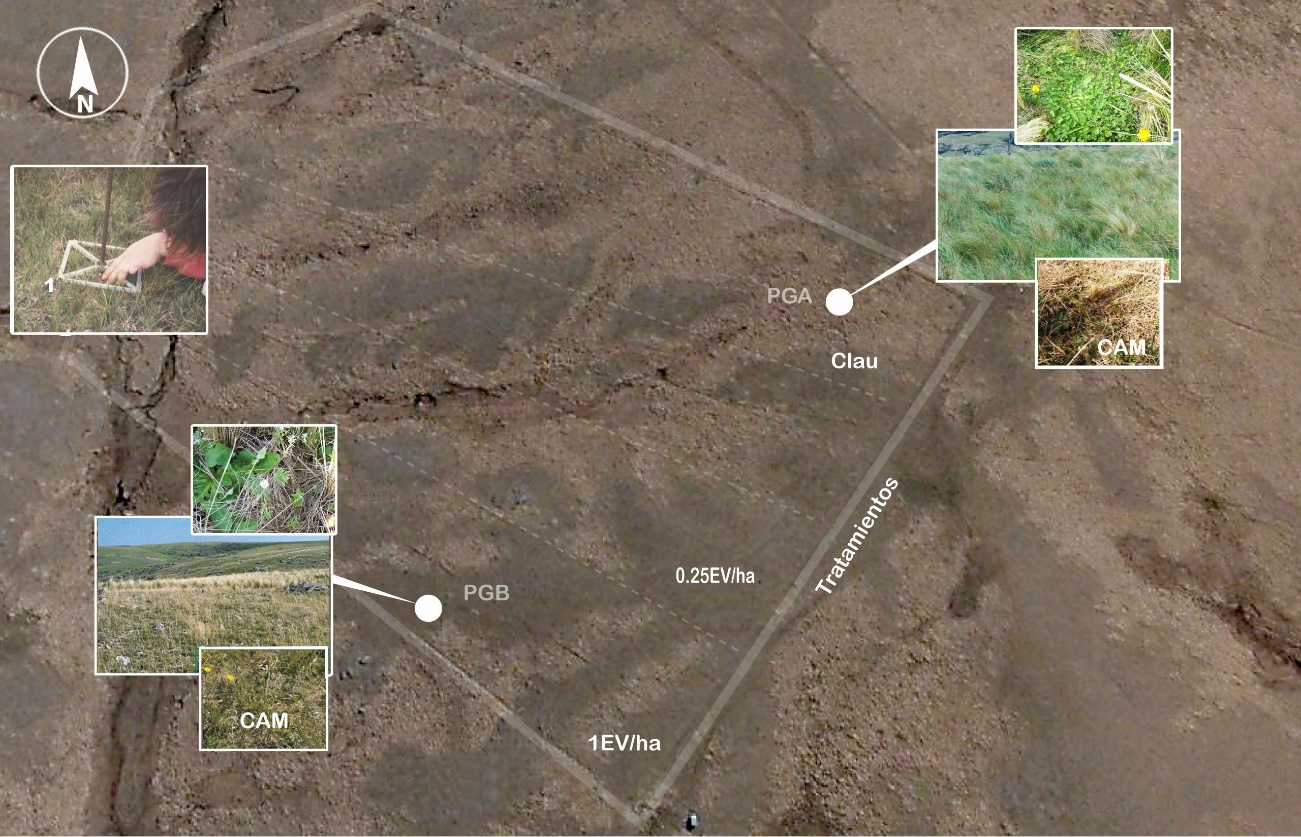 Figura 1
Detalle del ensayo
Figura 1
Detalle del ensayo
Abreviaturas: CAM (campo), PGA (pastizal de gramíneas altas), PGB (pastizal de gramíneas bajas). 1 parcela de lectura permanente, en 1EV/ha, 0,25 EV/ha y Clau (clausura)
Análisis de los datos
Las diferencias debidas al efecto del pastoreo sobre el tamaño del banco en cada comunidad fueron evaluadas a través de pruebas paramétricas (ANOVA de una vía y posteriormente Test de Tukey). Se utilizaron transformaciones de la √2 de los datos (Zar 1999). Para evaluar la composición florística del BSS en el campo se realizó un análisis de ordenación (DCA), dicho análisis se realizó mediante el programa estadístico PC-ORD (McCune y Mefford 2018). Luego se emplearon: el índice de diversidad (Shannon-Weaver 1949), la riqueza, la equidad.
Los análisis fueron realizados con el software InfoStat, (Di Rienzo et al., 2020) y el programa estadístico SPSS. Para la nomenclatura de las especies se siguió a Zuloaga et al., (1994), Zuloaga y Morrone (1996, 1999) y también se consultó el Catálogo on line de Las Plantas Vasculares de la Argentina, del Instituto de Botánica Darwinion (http://buscador.floraargentina.edu.ar/)
Resultados
Tamaño del BSS
Del análisis Tabla de ambos sitios se observó que no hubo diferencias (p< 0,05) en el tamaño del BSS entre los mismos (Tabla 1). Cuando se analizó el tamaño del BSS (Figura 2) en la combinación de los dos sitios con los tres tratamientos, se pudo observar que los tamaños de BSS fueron mayores para 1EV/ha, siguiéndole el de 0,25 EV/ha y el menor para la clausura, independientemente del sitio.
Tabla 1
Tamaño del BSS N° de semillas/m2 para PGA PGB
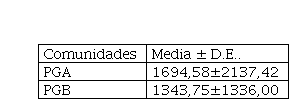
| Comunidades | Media ± D.E.. |
| PGA | 1694,58±2137,42 |
| PGB | 1343,75±1336,00 |
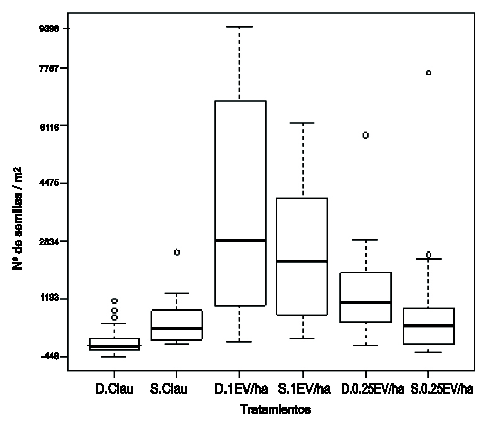 Figura 2
Diagrama de cajas correspondiente al tamaño del BSS (semillas/m2), para los diferentes tratamientos de pastoreo y clausura, en PGA (D) y PGB (S)
Figura 2
Diagrama de cajas correspondiente al tamaño del BSS (semillas/m2), para los diferentes tratamientos de pastoreo y clausura, en PGA (D) y PGB (S)
El ANOVA mostró que no hubo diferencias significativas entre sitios (p = 0.60303), mientras que sí se observaron entre tratamientos (p= 8.278e-12), y también en la interacción sitio y tratamiento, (p= 0.01725). En la Figura 3, se observó que el factor sitio tuvo un efecto diferente en la clausura con respecto a los otros tratamientos, ya que el tamaño de BSS para dicho tratamiento fue mayor en la comunidad de PGB que en la comunidad de PGA, lo que no ocurrió con los tratamientos pastoreados, donde el tamaño de BSS fue mayor para PGA. Al realizar un test a posteriori de comparación de medias se comprobó que hubo diferencias estadísticamente significativas (p < 0,05) entre los sitios para los tratamientos con pastoreo. En la comunidad de PGA, los tratamientos con pastoreo se diferencian significativamente de la clausura, en tanto que en PGB, es el pastoreo severo el que se diferencia de manera significativa (p < 0,05) del resto de los tratamientos.
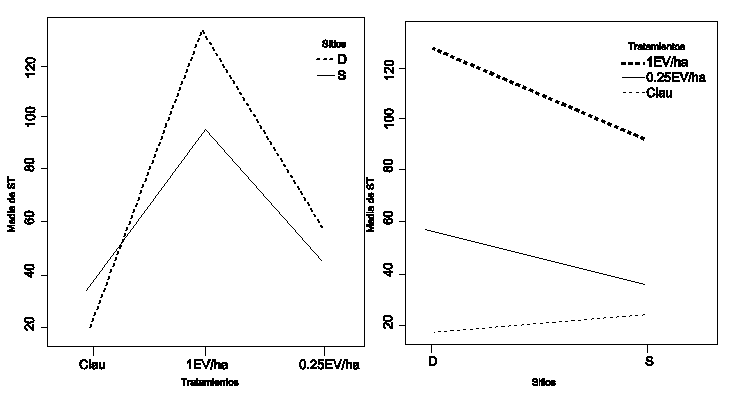 Figura 3
Interacción sitio-tratamientos (S: T) para (S) PGA y PGB
Figura 3
Interacción sitio-tratamientos (S: T) para (S) PGA y PGB
Nivel de significancia p<0.05.
El aporte de monocotiledóneas y dicotiledóneas al BSS en PGA fue mayor (p < 0,05) en el tratamiento con pastoreo severo; en PGB el tratamiento con pastoreo moderado mostró el aporte mayor de las monocotiledóneas, en cambio la contribución de las dicotiledóneas fue superior en el tratamiento con pastoreo severo, siendo las diferencias estadísticamente significativas.
En PGA y PGB, el tratamiento con pastoreo severo involucró la mayor contribución al BSS tanto para las especies anuales como para las perennes, siendo la diferencia estadísticamente significativa (p < 0,05) con respecto a los otros tratamientos.
Caracterización florística
Se relevaron un total de cincuenta y nueve especies, veintisiete fueron comunes en las comunidades estudiadas. En el pastizal alto, diez especies se presentaron de forma exclusiva, en tanto en el pastizal bajo fueron veintidós las especies exclusivas. En el análisis de ordenamiento de las comunidades (Figura 4a) se percibió como se diferencian los PGA y PGB en función de la composición florística, evidencia que fue explicada en gran magnitud por el eje 1: eigenvalue=0.79 y en menor medida por el eje 2: eigenvalue=0.50. Cuando se realizó el ordenamiento de las réplicas en cada comunidad en función de los tratamientos y la composición florística, se visualizaron mayores diferencias florísticas en PGA entre el tratamiento con pastoreo severo y la clausura, (Figura 4b, eje:2) y en PGB, entre los tratamientos pastoreados y la exclusión al mismo.
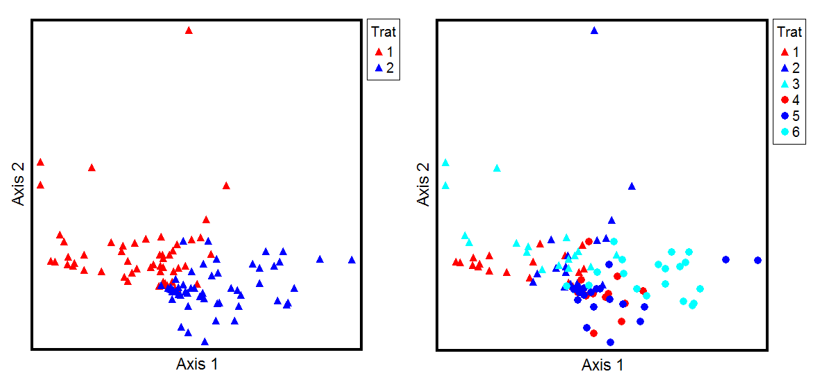 Figura 4
Figura 4
a) Ordenamiento de las réplicas considerando las comunidades, según eje 1 y eje 2 de DCA, PGA rojo y PGB azul. b) Ordenamiento de las réplicas del BSS considerando cada tratamiento, según eje 1 y eje 2 de DCA, (triángulos) PGA; (círculos) PGB; 1Ev/ha (rojo), 0,25EV/ha (azul) y Clausura (celeste).
Tabla 2
Riqueza (S) equidad (J´) y diversidad (H´) del BSS
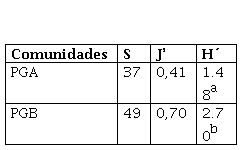
| Comunidades | S | J’ | H´ |
| PGA | 37 | 0,41 | 1.48a |
| PGB | 49 | 0,70 | 2.70b |
Se observaron diferencias estadísticamente significativas (p< 0,05), siendo mayor la riqueza, equidad y la diversidad en la comunidad de PGB (Tabla 2). Cuando se analizó riqueza, diversidad y equidad, dentro de cada una de las comunidades y tratamientos, se registraron diferencias significativas en PGA y PGB, siendo el tratamiento de pastoreo severo el que mostró los menores valores de diversidad y equidad, en tanto que la clausura registró el mayor valor para dichos descriptores, mientras que el valor de riqueza fue superior en el tratamiento con pastoreo moderado (Tabla 3 y 4).
Tabla 3
Riqueza (S), equidad (J´) y diversidad (H´) de las especies del BSS del PGA, en los diferentes tratamientos.
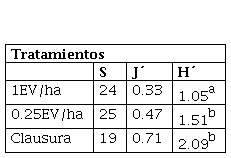
| Tratamientos |
| S | J´ | H´ |
| 1EV/ha | 24 | 0.33 | 1.05a |
| 0.25EV/ha | 25 | 0.47 | 1.51b |
| Clausura | 19 | 0.71 | 2.09b |
Tabla 4
Riqueza (S), equidad (J´) y diversidad (H´) de las especies del BSS del PGB, en los diferentes tratamientos.

| Tratamientos |
| S | J´ | H´ |
| 1EV/ha | 32 | 0.27 | 0.92b |
| 0.25EV/ha | 36 | 0.44 | 1.58b |
| Clausura | 32 | 0.66 | 2.3ª |
Se identificaron 5 especies exóticas en común entre PGB y PGA: Acicarpha tribuloides, Carduus thoermeri, Cirsium vulgare, Lolium multiflorumde crecimiento anual-invernal y Taraxacum officinale, de crecimiento perenne-invernal; además, en PGA se censaron otras tres especies exóticas: Lysimachia minima, Geranium dissectum de crecimiento anual-invernal y Trifolium repens, de crecimiento perenne-invernal.
En PGA dieciocho especies fueron exclusivas de los tratamientos con pastoreo y dieciséis lo fueron en la comunidad de PGB. En los sitios pastoreados se destacaron especies de buena calidad forrajera; en PGA se hallaron a Chascolytrum subaristatum (nativa) y Trifolium repens (exótica); en PGB ocurrió solo una, Sorghastrum pellitum (nativa). Como especie invasora se relevó a Carduus thoermeri en ambas comunidades.
Dentro de las especies con mayor tamaño de BSS en los sitios pastoreados, Hypochaeris chillensis fue común en las dos comunidades, mientras que en PGA lo fueron: Lolium multiflorum, Hypoxis humilis . Taraxacum officinale y en PGB: Bulbostylis juncoides, Arenaria achalensis. Muhlenbergia peruviana. En el tratamiento con exclusión al pastoreo, las especies de mayor contribución al BSS fueron en la comunidad de PGA: Bromus catharticus, Lolium multiflorum y Taraxacum officinale y en la comunidad de PGB: Hypochaeris chillensis, Nassella filiculmis, Muhlenbergia peruviana.
En ambas comunidades se censaron especies con escasa contribución al BSS, pero que son importantes por sus características de exóticas e invasoras: Cirsium vulgare, Conyza bonariensis, Carduus thoermeri . Taraxacum officinale; además Nassella filiculmis considerada una invasora-nativa. En cambio, dentro de las especies comunes, se encontraron algunas importantes por su contribución al BSS, como Lolium multiflorum e Hypochaeris chillensis; para PGA, en tanto que en PGB la especie de mayor contribución fue Hypochaeris chillensis (nativa e invasora). Otra especie que se registró en las dos comunidades y que es importante por su buena calidad forrajera fue Sorghastrum pellitum. Especies con la misma importancia forrajera registrada solo en PGA fueron Bromus auleticus, y Trifolium repens, mientras que solo en PGB se observó a Lolium multiflorum, sin embargo, no estuvo presente en el tratamiento con pastoreo severo.
Discusión
Al analizar la composición florística, las comunidades compartieron la mitad de las especies relevadas en el BSS. En ambas comunidades, los tratamientos pastoreados y la clausura evidenciaron diferencias en la composición florística. Las dos comunidades poseen estructuras diferentes (Cantero et al., 2001) y reaccionan en forma desigual frente a la presión de pastoreo. El pastoreo simplifica el estrato herbáceo permitiendo que especies arrosetadas sean las especies de mayor emergencia aprovechando la apertura del canopeo y los sitios seguros para germinar en ambas comunidades y en el caso de PGB, pueden llegar a reemplazar a especies palatables (Westoby y Wright, 2006 y Rauber et al., 2023). Los cambios provocados por el pastoreo en pastizales nativos en todo el mundo ya han sido reportados por numerosos autores (Milchunas y Laurenroth, 1993, Cingolani et al., 2008).
Los valores de riqueza, equidad y diversidad fueron significativamente mayores en PGB en relación a PGA, coincidiendo con lo reportado por Funes et al., (2001) en experimentos realizados en invernáculo. La diferencia se debió principalmente a que en PGB germinaron muchas especies en roseta, postradas o graminoides, que aprovecharon la ventaja de los sitios de escasa cobertura o denudados (Loydi et al., 2012), generados por perturbación y que no podrían germinar bajo condiciones tales como presencia de alta cantidad de mantillo (Baskin y Baskin, 2014), situación que se da en PGA.
En cuanto a los tratamientos, en ambas comunidades, con la exclusión al pastoreo ocurrieron los mayores valores de diversidad y de equidad, mientras que con el pastoreo severo se observaron los menores valor de diversidad y de equidad, lo cual concuerda con lo expresado por Cingolani et al., (2005); Rauber et al., (2023) quienes sostiene que, ante perturbaciones severas, la diversidad disminuye.
Se reportaron la presencia de especies exóticas en las dos comunidades y en los tratamientos pastoreados y con exclusión, siendo Carduus thoermeri, Cirsium vulgare . Taraxacum officinale las más importantes y con mayor incidencia en el tratamiento con pastoreo severo. Estos resultados no coinciden con lo reportado Márquez et al., (2002), quienes encontraron una baja cantidad de especies exóticas, sugiriendo que los pastoreos a intensidades moderadas a altas no estarían actuando como una perturbación severa. Sin embargo, Amuchástegui (2016), no registró un aumento de especies exóticas en el BSS, pero sí un aumento en su contribución al mismo en los sitios pastoreados en forma severa, lo cual podría indicar un deterioro del pastizal sí persistiera dicho pastoreo, en concordancia con lo reportado por Vecchio et al., (2022).
El tamaño del BSS no mostró diferencias significativas entre las dos comunidades. En cuanto a los tratamientos, se observó una tendencia similar en donde los mayores tamaños estuvieron en los tratamientos con pastoreo severo seguido por el pastoreo moderado y por último la clausura (Minervini Silva, 2020). Los tamaños de BSS fueron mayores en PGA en relación a PGB en los tratamientos pastoreados, diferencia que fue estadísticamente significativa, mientras que la clausura de PGB mostró el comportamiento inverso. En el tratamiento con exclusión al pastoreo en PGA se genera una mayor acumulación de mantillo que impide que las semillas tomen contacto con el suelo (Márquez et al., 2002), disminuyendo el enterramiento de las mismas y dejándolas expuestas a la predación, sumado a la mayor cobertura de la vegetación establecida que disminuye la llegada de la luz incidente, pudiendo provocar la dormancia en algunas especies (Baskin y Baskin, 2014).
En PGB el pastoreo moderado no se diferenció estadísticamente de la clausura en el tamaño de BSS, debido a que esta comunidad es más tolerante al pastoreo (Nuñez et al., 2001). Aquí es probable que el pastoreo moderado no sea una perturbación que llegue a producir cambios importantes en cuanto al tamaño de BSS. Estos resultados coinciden con los hallados en otros pastizales que resisten bien el efecto del pastoreo moderado (Cingolani et al., 2015).
En ambas comunidades quedó demostrado que el pastoreo severo se diferencia en cuanto a tamaño del BSS, del tratamiento con exclusión al pastoreo. Estos resultados fueron coincidentes con lo reportado por Marco y Páez, (2000) pero no coincidieron con Tessema et al., (2016) y Loydi et al., (2019). Cabe aclarar que estos estudios fueron realizados en invernáculo, ya que no se han encontrado trabajos publicados en donde los censos hayan sido realizados directamente a campo.
Sandoval–Calderón et al., 2024, coinciden con lo reportado aquí, en ambas comunidades con respecto al efecto del pastoreo severo en el banco de semillas. Estos autores expresan que ante el pastoreo con altas cargas, se provocan efectos diferenciales en la dispersión, establecimiento y sobrevivencia entre las especies palatables y las que no lo son, ya que el pastoreo selectivo produce cambios en las jerarquías competitivas (Kahmen et al, 2002,), proceso que se ve facilitado por la disminución del área foliar de las especies dominantes (Kahmen y Poschlod, 2008), lo que facilitaría el establecimiento y posterior reemplazo de las mismas por otras especies menos palatables.
Las especies que se vieron favorecidas en PGA, fueron: Lolium multiflorum Hypochaeris chillensis. En estos pastizales, Hypochaeris chillensis es una de las más eficientes en la colonización de micrositios en ambas comunidades (Amuchástegui, 2016), debido a que el pastoreo severo reduce la cobertura vegetal y genera mayor cantidad de sitios seguros para la germinación y establecimiento de dicha especie, la cual invade el pastizal, desmejorando su condición para el pastoreo.
En ambas comunidades el BSS estuvo representado en su mayor proporción por especies de escaso valor forrajero y otras oportunistas que poseen la capacidad de germinar en diversas situaciones (Taraxacum officinale, Hypochaeris chillensis . Muhlenbergia peruviana). Loydi et al., 2012, reportó resultados similares en cuanto a que las especies palatables aparecen esporádicamente en el BSS.
En las dos comunidades Hypochaeris chillensisfue la especie en común de mayor aporte al tamaño del BSS. Esta especie tiene su mayor pulso de germinación en primavera, si bien también germina en otoño, ya que cumple rápidamente su ciclo de crecimiento y en los meses que se produce el pastoreo escapa al mismo. Puede aprovechar los descansos estacionales que se realizan en estos pastizales que van desde mediados de mayo a septiembre y con estas ventajas competitivas puede convertirse en una potencial invasora del pastizal (Amuchastegui, 2016). Estos resultados concuerdan con los resultados hallados Kahmen et al., (2002) quienes plantean que, en sitios pastoreados por altas cargas, la gran densidad de semillas de las dicotiledóneas presentes en el banco está relacionada con la presencia en la vegetación de dicotiledóneas no palatables de hábito rastrero y arrosetado, que se ven favorecidos por el pastoreo selectivo.
Conclusiones
El estudio comparativo de las comunidades de pastos altos (PGA) y pastos bajos (PGB) reveló que no existen diferencias significativas en el tamaño del banco de semillas del suelo (BSS) entre ambas comunidades. Sin embargo, los pastos altos mostraron mayores tamaños en los tratamientos pastoreados en comparación con los pastos bajos. Lo que pone en evidencia que, el pastoreo influenció significativamente en el tamaño del BSS. El pastoreo severo presentó los mayores valores, en ambas comunidades, cuya mayor contribución estuvo dada por un incremento notable de especies dicotiledóneas, especialmente Hypochaeris chillensis. La riqueza, equidad y diversidad fueron mayores en los pastos bajos y entre los tratamientos fue la exclusión al pastoreo el que resultó con los mayores valores de riqueza y diversidad para ambas comunidades. El pastoreo también incrementó las especies anuales monocotiledóneas y dicotiledóneas, con un mayor incremento de estas últimas en pastoreo severo. En dicho tratamiento, también fueron más prevalentes, las especies exóticas, destacándose Lolium multiflorum, Trifolium repens, Taraxacum officinale, Cirsium vulgare y Carduus thoermeri. Se concluye que las diferentes intensidades de pastoreo o su exclusión impactaron de manera diferencial en las comunidades vegetales de los pastizales serranos centro argentinos. La información generada sobre el BSS es fundamental para prescribir normas de manejo que favorezcan la conservación y los servicios ambientales de estos pastizales. En este sentido, para un manejo efectivo que concilie la producción con la conservación de la biodiversidad se recomienda ampliar estudios a largo plazo considerando la dinámica de los nutrientes, compactación del suelo y acumulación de broza, así como la relación entre germinación de semillas y establecimiento de la vegetación.
Agradecimientos
Secretaría de Ciencia y Tecnología de la Universidad Nacional de Río Cuarto (SECyT-UNRC) y a los productores de la zona que sin su apoyo no se hubiese podido realizar este experimento.
Referencias bibliográficas
Amuchástegui, M. A. (2016). Efectos del pastoreo sobre el banco de semillas del pastizal natural serrano [Tesis de Maestría, Universidad Nacional de Río Cuarto]. Escuela de Postgrado. (Inédito).
Bakker, J. P., Bos, A. F., Hoogveld, J., y Muller, H. J. (1991). The role of the seed bank in restoration management of semi-natural grasslands. In O. Ravera (Ed.), Terrestrial and aquatic ecosystems: Perturbation and recovery (pp. 449-455). Ellis Horwood.
Bakker, J. P., Poschlod, P., Strykstra, R. J., Bekker, R. M., y Thompson, K. (1996). Seed banks and seed dispersal: Important topics in restoration ecology. Acta Botanica Neerlandica, 45(4), 461-490.
Baskin, C. C., y Baskin, J. M. (2014). Seeds: Ecology, biogeography, and evolution of dormancy and germination (2nd ed.). Academic Press.
Cabrera, A. (1976). Regiones fitogeográficas argentinas. Enciclopedia Argentina de Agricultura y Jardinería. Tomo II, Fasc. I. ACME.
Cantero, J. J., y Bianco, C. A. (1986). Las plantas vasculares del suroeste de la Provincia de Córdoba. Parte III. Catálogo preliminar de especies. Revista UNRC, 6(1), 5-52.
Cantero, J. J., Pärtel, M., y Zobel, M. (1999). Is species richness dependent on the neighbouring stands? An analysis of the community patterns in mountain grasslands of central Argentina. Oikos, 87, 346-354.
Cantero, J. J., Cabido, M., Nuñez, C. O., Petryna, L., Zak, M. R., y Zobel, M. (2001). Clasificación de los pastizales de suelos sobre rocas metamórficas de las Sierras de Córdoba, Argentina. Kurtziana, 29(1), 27-77.
Catálogo de las Plantas Vasculares de la Argentina, Instituto de Botánica Darwinion. (s. f.). http://www.darwin.edu.ar/Proyectos/FloraArgentina/FA.asp
Cingolani, A. M., Noy-Meir, I., y Díaz, S. (2005). Grazing effects on rangeland diversity: A synthesis of contemporary models. Ecological Applications, 15(2), 757-773.
Cingolani, A. M., Noy-Meir, I., Renison, D. D., y Cabido, M. (2008). La ganadería extensiva: ¿es compatible con la conservación de la biodiversidad y de los suelos? Ecología Austral, 18(3), 253-271.
Cingolani, A. M., Vaieretti, M. V., Giorgisa, M. A., Poca, M., Tecco, P. A., y Gurvich, D. E. (2014). Can livestock grazing maintain landscape diversity and stability in an ecosystem that evolved with wild herbivores? Perspectives in Plant Ecology, Evolution and Systematics, 16, 143-153.
Coop, I. E. (1965). A review of the Ewe Equivalent System. New Zealand Agricultural Science, 1(3), 13-18.
Díaz, S., Hodgson, J. G., Thompson, K., y Cabido, M. (2004). The plant traits that drive ecosystems: Evidence from three continents. Journal of Vegetation Science, 15, 295-304.
Díaz, S., Fargione, J., Chapin, F. S., III, y Tilman, D. (2006). Biodiversity loss threatens human well-being. PLoSBiology, 4(8), e277. https://doi.org/10.1371/journal.pbio.0040277
Di Rienzo, J. A., Casanoves, F., Balzarini, M. G., Gonzalez, L., Tablada, M., y Robledo, C. W. (2017). InfoStatversión 2020. Grupo InfoStat, FCA, Universidad Nacional de Córdoba. http://www.infostat.com
Facelli, J. M., y Facelli, E. (1993). Interactions after death: Plant litter controls priority effects in a successional plant community. Oecologia, 95, 277-282.
Fenner, M., y Thompson, K. (2005). The ecology of seeds. Cambridge University.
Funes, G., y Cabido, M. (1995). Variabilidad local y regional de la vegetación rupícola de las Sierras Grandes de Córdoba, Argentina. Kurtziana, 24, 173-188.
Funes, G., Basconcelos, S., Díaz, S., y Cabido, M. (2001). Edaphic patchiness influences grassland regeneration from the soil seed bank in mountain grasslands of central Argentina. Austral Ecology, 26, 205-212.
Gonzalez, J., Cantero, J. J., Nuñez, C. O., Cholaky, C., Cisneros, J., y Petryna, L. (1999). Cartografía de los ecogrupos vegetales en los paisajes serranos del centro sur de Córdoba, Argentina. Revista UNRC, 19(1-2), 5-17.
Haretche, F., y Rodríguez, C. (2006). Banco de semillas de un pastizal uruguayo bajo diferentes condiciones de pastoreo. Ecología Austral, 16, 105-113.
Kahmen, S., y Poschlod, P. (2008). Effects of grassland management on plant functional trait composition. Agriculture, Ecosystems y Environment, 128(3), 137-145. https://doi.org/10.1016/j.agee.2008.05.016
Kahmen, S., Poschlod, P., y Schreiber, K. F. (2002). Conservation management of calcareous grasslands. Changes in plant species composition and response of functional traits during 25 years. Biological Conservation, 104, 319-328.
Lipoma, M. L., Funes, G., y Díaz, S. (2018). Fire effects on the soil seed bank and post-fire resilience of a semi-arid shrubland in central Argentina. Austral Ecology, 43, 46-55.
Loydi, A., Zalba, S. M., y Distel, R. A. (2012). Viable seed banks under grazing and exclosure conditions in montane mesic grasslands of Argentina. Acta Oecologica, 43, 8-15.
Loydi, A. (2019). Effects of grazing exclusion on vegetation and seed bank composition in a mesic mountain grassland in Argentina. Plant Ecology and Diversity, 12(2), 127-138.
Luti, R., Solís, A., Galera, F. M., Müller, N., Berzal, M., Nores, M., Herrera, M., y Barrera, J. C. (1979). Vegetación. Geografía Física de la Provincia de Córdoba. In J. B. Vázquez, R. A. Miatello, y M. E. Roqué (Eds.). Buenos Aires: Boldt.
McCune, B., y Mefford, M. J. (2018). PC-ORD: Multivariate Analysis of Ecological Data (Version 7). MjM Software.
Marberg, M. (2013). Climate, grazing and plant interactions: Does climate and grazing shape plant interactions in alpine environments? [Degree Thesis in Biology]. Supervisor: Johan Olofsson.
Marco, D. E., y Páez, S. A. (2000). Soil seed banks on Argentine seminatural mountain grasslands after cessation of grazing. Mountain Research and Development, 20(3), 254-261.
Márquez, S., Funes, G., Cabido, M., y Pucheta, E. (2002). Efectos del pastoreo sobre el banco de semillas germinable y la vegetación establecida en pastizales de montaña del centro de Argentina. Revista Chilena de Historia Natural, 75, 327-337.
Milberg, P. (1995). Soil seed bank after eighteen years of succession from grassland to forest. Oikos, 72, 3-13.
Milchunas, D. G., y Lauenroth, W. K. (1993). A quantitative assessment of the effects of grazing on vegetation and soil over a global range of environments. Ecological Monographs, 63, 327-366.
Minervini Silva, G., y Overbeck, G. E. (2020). Soil seed bank in a subtropical grassland under different grazing intensities. Acta Botanica Brasilica, 34(2), 360-370.
Morici, E., Doménech-García, V., Dorado-Rodríguez, R., y Gallart, F. (2017). Trophic interactions in a Patagonian dry steppe: Effects of sheep grazing on soil seed banks and vegetation. Basic and Applied Ecology, 24, 42-51.
Müller, I., y Böhning-Gaese, K. (2003). Seed bank composition of semi-natural grasslands in relation to grazing. Journal of Vegetation Science, 14, 319-328.
Peco, B., Traba, J., Levassor, C., Sánchez, A. M., y Azcárate, F. M. (2003). Seed size, shape and persistence in dry Mediterranean grass and scrublands. Journal of Vegetation Science, 14, 335-342.
Peco, B., Ortega, M., y Levassor, C. (1998). Similarity between seed bank and vegetation in Mediterranean grassland: A predictive model. Journal of Vegetation Science, 9(6), 815-828.
Peña, M. (2004). Contribución al conocimiento del banco de semillas de especies herbáceas de la Pampa Ondulada. Tesis de Licenciatura, Universidad Nacional de Luján. (Inédito).
Prieto, I., Echeverría, E., Funes, G., y Cabido, M. (2002). Diversidad florística de pastizales de suelos sobre rocas ultrabásicas en las Sierras de Córdoba, Argentina. Ecología Austral, 12, 59-74.
Pyke, D. A. (1986). Demographic responses of Bromus tectorum and seedlings to grazing by small mammals: the influence of grazing and dispersal pattern. Oecologia, 70, 7-16.
Pywell, R. F., Bullock, J. M., Walker, K. J., Coulson, S. J., Gregory, S. J., y Stevenson, M. J. (2004). Facilitating grassland diversification using the soil seed bank: an experimental assessment of multiple techniques. Biological Conservation, 119, 537-548.
Rusch, G. M. (1992). Legumes in seed banks in grazed and ungrazed northern hill grasslands. Oikos, 63(2), 172-181.
Ryser, P., y Lamberti-Raverot, B. (1992). Plant functional types and soil seed banks: The experience of a calcareous grassland. Journal of Vegetation Science, 3, 65-74.
Sala, O. E., Golluscio, R. A., Lauenroth, W. K., y Soriano, A. (1989). Resource partitioning between shrubs and grasses in Patagonia: A hypothesis based on vegetation isotopic composition. Acta Oecologica, 10(1), 21-30.
Schreiber, K. F. (Ed.). (1987). Restoration ecology: A synthetic approach to ecological research. Cambridge University.
Smith, R. S., y Jones, L. (2002). The role of the seed bank in restoration management of wet grasslands. Journal of Applied Ecology, 39, 464-476.
Smith, R. S., Shiel, R. S., Millward, D., y Younger, A. (2008). Long-term effects of inorganic fertilizers and lime on the seed bank of upland grassland. Agriculture, Ecosystems y Environment, 128, 99-106.
Thompson, K., Bakker, J. P., y Bekker, R. M. (1997). The soil seed banks of north west Europe: Methodology, density and longevity. Cambridge University.
Willems, J. H. (2001). Problems, approaches, and results in restoration of Dutch calcareous grassland during the last 30 years. Restoration Ecology, 9(2), 147-154.
Wu, Z. H., Yang, Y. F., y Hu, Z. Y. (2006). Effects of grazing on soil seed banks in temperate steppe of Inner Mongolia. Acta Ecologica Sinica, 26(10), 3351-3359.
Young, J. A., y Young, C. G. (1992). Seeds of woody plants in North America. Dioscorides Press.
Zar, J. H. (1999). Biostatistical analysis (4th ed.). Prentice Hall.
Zuloaga, F. O., y Morrone, O. (1996). Catálogo de las plantas vasculares de la República Argentina. I. Pteridophyta, Gymnospermae y Angiospermae (Monocotyledoneae). Monographs in Systematic Botany from the Missouri Botanical Garden (Vol. 60).
Zuloaga, F. O., y Morrone, O. (1999). Catálogo de las plantas vasculares de la República Argentina. II. Dicotyledoneae. Monographs in Systematic Botany from the Missouri Botanical Garden (Vol. 74).
Zuloaga, F. O., Nicora, E. G., Rúgolo de Agrasar, Z. E., Morrone, O., Pensiero, J., y Cialdella, A. M. (1994). Catálogo de la familia Poaceae en la República Argentina. Monographsin Systematic Botany from the Missouri Botanical Garden (Vol. 47).
Notas de autor
aamuchastegui@ayv.unrc.edu.ar





