Resumen: La invención de la electroencefalografía (EEG) marcó un hito histórico tecnológico/metodológico que posicionó a la actividad eléctrica cerebral de gran escala como nuevo objeto epistemológico para el estudio de fenómenos cognitivos, inaugurando la subdisciplina de la Electrofisiología Cognitiva. Este documenta una historia breve de la Electrofisiología Cognitiva, desde los orígenes de la Electrofisiología Cognitiva, su apogeo científico/social en la década del 1980, hasta la invención de la resonancia magnética funcional. Posteriormente, revisamos aspectos metodológicos y teóricos predominantes en la literatura respecto al potencial relacionado a eventos (event related potential, ERP). Además, se provee software para computar ERP y datos de muestra. Actualmente, las señales neurofisiológicas humanas adquiridas no-invasivamente siguen siendo una de las principales formas de estudiar correlatos neurales de fenómenos cognitivos. El presente trabajo busca promover las herramientas necesarias para el entendimiento sobre la computación e interpretación de estas señales cerebrales.
Palabras clave: EEG, Electrofisiología Cognitiva, Potenciales Relacionados a Eventos, Software,Metodología.
Abstract: Electroencephalography (EEG) invention set a historic technological/methodological milestone, positioning large-scale brain electrical activity as a new epistemological object for the study of cognitive phenomena, promptly inaugurating the Cognitive Electrophysiology subdiscipline This manuscript documents a brief history of Cognitive Electrophysiology, the historical account of the development of Cognitive Electrophysiology from its origins and its scientific/societal apogee in the 1980’s, to the invention of functional magnetic resonance imaging (fMRI) Then, we proceed to review methodological and theoretical guidelines of the most widely used technique in cognitive electrophysiology, event-related potentials (ERP). We also provide software that allows computing ERPs with the described techniques along with sample data. Nowadays, non-invasively acquired neurophysiological signals continue to be one of the main ways to study neural correlates of cognitive phenomena. The present work promotes necessary tools for an accurate understanding of computing and interpreting brain signals.
Keywords: EEG, Cognitive Electrophysiology, Event- Related Potentials, Software, Methodology.
Computación, análisis e interpretación de señales eléctricas del cerebro humano en el dominio del tiempo: 90 años de Electrofisiología Cognitiva
Autor de correspondencia: francisco.parada@udp.cl
El reporte de actividad cerebral eléctrica adquirida no-invasivamente desde un cerebro humano en vivo cumplió 90 años (Berger, 1929). Los estudios neurocientíficos actuales buscan entender procesos neurales y mentales subyacentes a la experiencia perceptiva del observador enfrentado a eventos y/o a la ejecución de una respuesta o decisión frente a los mismos usando electroencefalograma (EEG) (Krause et al., 1996; Pfurtscheller & da Silva, 1999). Asimismo, EEG es utilizado para informar -implícitamente- acerca del estado mental de un individuo, ya sea este de alerta, somnolencia o anestesia (Nakamura et al., 1996; Purdon et al., 2013). De esta manera, EEG es una herramienta que sirve tanto para diagnóstico clínico como para investigación en ciencias psicológicas (Biasiucci et al., 2019; Cohen, 2014; Niedermeyer & da Silva, 2005; Tong & Thakor, 2009).
Sin embargo, a 90 años de su invención, aún existe muy poca congruencia entre teoría y práctica respecto a su uso. Esta desorganización conceptual (Cohen, 2014) ha dificultado que estudiantes e investigadores provenientes de la Psicología y las Ciencias Sociales puedan dimensionar el impacto epistemológico/metodológico que la Electrofisiología Cognitiva puede añadir a sus agendas de investigación.
El presente trabajo busca (i) documentar una historia breve de la Electrofisiología Cognitiva, (ii) proveer lineamientos teóricos y prácticos para la computación, análisis e interpretación de señales neurales en el dominio del tiempo y (iii) entregar software para MATLAB (The Mathworks Inc., Natick, MA) que permite implementar las prácticas descritas.
El EEG contemporáneo es el resultado de un esfuerzo científico teórico/tecnológico que se ha extendido por más de 200 años. En una disciplina con tan larga trayectoria como la Neurociencia, conocer el contexto histórico que da origen a nuestras prácticas es esencial, puesto que permite tener una mirada crítica al momento de revisar, analizar e interpretar datos. Es por esto que antes de entender la técnica de análisis/interpretación de señales neurales, queremos proveer un breve recorrido histórico.
En la última década del siglo XVIII, el médico y filósofo Luigi Galvani se desempeñó como anatomista teórico en la Academia de Ciencias de Boloña. Fuertemente influenciado por el trabajo de Jan Swammerdam[1] (Stillings, 1975), Galvani realizó experimentos donde vinculó electricidad estática y tejido vivo. Trabajos posteriores establecen la relación entre flujos eléctricos y el funcionamiento de fibras musculares (Piccolino, 1998). De esta manera, la bioelectricidad posiciona a los fenómenos eléctricos que ocurren en nervios y músculos como un objeto de estudio científico.
Casi 50 años más tarde, Emil Du Bois-Reymond propuso que los fenómenos observados por Galvani y otros se deben a cambios discretos, rápidos y transitorios de actividad eléctrica. Du Bois-Reymond (1848) los denominó como “corrientes de acción nerviosa”. Este descubrimiento, sumado a los posteriores aportes de J. Bernstein, K.S. Cole, H. J. Curtis, A. Hodgkin y A. Huxley, dieron paso a lo que actualmente se conoce como potencial de acción; la unidad dinámica básica del operar del sistema nervioso (Collura, 1993).
La relevancia teórica del potencial de acción reside en el entendendimiento de cada neurona como un dipolo eléctrico[2]. Durante la propagación del potencial de acción, el interior de la membrana adquiere una carga positiva mientras que su exterior se torna negativo[3]. Cuando una corriente de despolarización logra abrir canales de sodio voltaje-dependientes, un potencial de acción se propaga desde las dendritas al soma y luego al axón (Kandel et al., 2000). Con el descubrimiento del potencial de acción se origina la electrofisiología.
El siglo XIX termina con la aceptación científica de que actividad eléctrica está a la base del operar del cuerpo de invertebrados, reptiles y mamíferos pequeños[4]. Fue difícil aceptar para la sociedad de aquel entonces que el cuerpo humano se componía de tan simples mecanismos. Esta idea comienza a cambiar en 1875, cuando Richard Caton registra impulsos eléctricos en tejido nervioso mamífero de conejos y monos. Trabajos posteriores demuestran que actividad bioeléctrica ocurre en el tejido nervioso periférico y central de todos los seres vivos, incluidos mamíferos y primates (Duffy et al., 1989).
A principios del siglo XX se registran potenciales bioeléctricos de manera no-invasiva en animales vivos. En 1929 el neurólogo y psiquiatra Hans Berger reportó, por primera vez, el registro electroencefalográfico de actividad cerebral en humanos (Berger, 1929). En el seminal artículo de 1929 se describe actividad cerebral alrededor de los 10 Hz, lo que en ese entonces se le llamó "el ritmo de Berger". Estas señales mostraban ciclos constantes y de mayor amplitud mientras la persona permanecía despierta pero con los ojos cerrados. En contraste, la amplitud y ritmicidad de las señales decrecía cuando las personas abrían sus ojos[5]. En 1939 el primer Atlas de Electroencefalografía sistematiza el trabajo realizado durante la primera década del EEG (Gibbs & Gibbs, 1939). Posteriormente, las señales eléctricas registradas por medio de electrodos de superficie son aceptadas en comunidades científicas como producto de actividad post-sináptica[6] de múltiples fuentes cerebrales (>50.000 unidades), sumada linealmente en el cuero cabelludo[7] (Nunez & Srinivasan, 2006).
EEG establece el primer vínculo entre estados perceptuales/cognitivos y dinámicas eléctricas cerebrales. Además, la elegante tarea conductual ideada por Berger es el primer registro de actividad cerebral en reposo, una técnica de suma relevancia para la neurociencia moderna (resting-state activity, revisada por Rosazza & Minati, 2011). Esta innovación tecnológica/metodológica genera un nuevo objeto epistemológico y con él, un nicho interdisciplinario que busca entender la relación entre dinámicas eléctricas del cerebro y la diversidad de estados mentales: la Electrofisiología Cognitiva. Posteriores avances e innovaciones tecnológico/metodológicas entregaron a la nueva disciplina una herramienta que la consolidó durante la década de 1980.
Estudios cualitativos caracterizaban la naciente disciplina de la electrofisiología cognitiva, sin embargo, el uso de valores centrales -o más típicos- para describir las principales tendencias neurodinámicas observadas luego de la percepción de estímulos sensoriales marcó el inicio de una revolución científica. El primer reporte de este tipo se hizo utilizando estimulación auditiva en participantes dormidos (Davis et al., 1939). Cabe recordar que estos análisis son previos a la invención del computador, por lo que el cálculo del ERP se implementó de forma análoga (Dawson, 1947) y -posteriormente- de manera electrónica (Dawson, 1954). No fue hasta la década de 1960 que se reporta el primer promedio de señales EEG calculado computacionalmente (Galambos & Sheatz, 1962). Este descriptor estadístico cuantitativo aumentó la precisión temporal con la que se reportan cambios en señales cerebrales. Asimismo, la inclusión de computadores en el análisis del EEG marcó el inicio de sus "años dorados". De esta manera, los potenciales relacionados a eventos (event-related potential, ERP) se establecen como la técnica de análisis estándar y de referencia para el estudio de fenómenos perceptuales y cognitivos.
En los siguientes 30 años, diversos científicos de la cognición utilizaron ERP para estudiar correlatos neurales de los fenómenos mentales (Donchin, 1979; Luck, 2014; van Beijsterveldt & Boomsma, 1994). El trabajo desarrollado por estos científicos cimentó al cerebro como un objeto epistemológico de relevancia social, tanto así que E.E.U.U. declaró los años 90 como “la década del cerebro” (Jones & Mendell, 1999). Esta década culminó con varios avances tecnológicos que permitían estudiar estructuras cerebrales y su función metabólica por medio de la Tomografía por Emisión de Positrones (Positron-Emission Tomography, PET Hoffman et al., 1976; Phelps et al., 1976; Posner et al., 1988; Sweet, 1951). Posteriormente, un nuevo método de imagenología aparecía: la Resonancia Magnética Funcional (functional magnetic resonance imaging, fMRI Bandettini et al., 1992). El fMRI se caracteriza por poseer una alta resolución espacial, lo cual permite el acceso a la estructura cerebral. Con el advenimiento del fMRI se evidenció una pérdida de la posición privilegiada de EEG dentro de la imagenología cerebral funcional, sin embargo, ERP sigue siendo una técnica robusta y con amplio respaldo en la literatura. A continuación, profundizaremos en su computación, orígenes fisiológicos e interpretación funcional.
Actualmente, la neurociencia posee una gran diversidad de métodos y técnicas para poder cuantificar la actividad cerebral (Senior et al., 2006). Sin embargo, los ERP siguen siendo la técnica de análisis utilizada con mayor frecuencia al momento de estudiar dinámicas cerebrales registradas en el cuero cabelludo. El éxito de esta técnica se debe, en parte, a su sencillez computacional y -aparentemente simple- interpretación funcional.
Luego de identificar datos que posean señales no-cerebrales de carácter no-lineal, los investigadores remueven artefactos predecibles/periódicos (por ej. movimientos oculares) con técnicas computacionales (por ej. machine learning) (Makeig & Onton, 2009). Tradicionalmente, filtros digitales de pasa-baja (lowpass) entre 25 y 40 Hz se aplican a los datos (Widmann et al., 2015), sin embargo, recientemente esta práctica se ha puesto en duda dada la gran cantidad de artefactos que pueden ser introducidos en la señal (Vanrullen, 2011; Widmann et al., 2015). Posteriormente, los datos se normalizan al sustraer actividad cerebral registrada antes de la aparición del estímulo a la actividad de interés, es decir, después de la aparición del estímulo. La normalización se realiza ensayo por ensayo (Grandchamp & Delorme, 2011). Así, los ERP se derivan a partir del promedio aritmético de un número suficiente de ensayos experimentales de actividad cerebral. Ecuación 1 fue implementada en la función computacional CalculaERP.m y declara este procedimiento, donde el ERP en un sensor es estimado a partir de la señal (x) registrada a través de un número (n) de ensayos experimentales (trials).
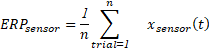 [Ecuación 1 - computación del ERP]
[Ecuación 1 - computación del ERP]Los ERP describen desviaciones promedio respecto a la actividad cerebral observada antes de la estimulación sensorial (i.e. línea base). Usualmente se describen en microvolts (µV) y poseen alta precisión temporal en el rango de los milisegundos (ms) lo que los convierte en idóneos testigos de los dinámicos fenómenos perceptuales y cognitivos. La señal ERP se define como la diferencia en fluctuaciones de potencial eléctrico cuyas dinámicas temporales y de fase permanecen fijas en relación a un evento físico, cognitivo y/o fisiológico determinado. Debido a que la electricidad se propaga instantáneamente a través de la superficie del cuero cabelludo, el origen exacto de esas señales sólo se puede estimar teóricamente (Cohen, 2014; Nunez & Srinivasan, 2006). Así, la principal desventaja del EEG/ERP es su baja resolución espacial, impidiendo responder con certeza preguntas acerca de neuroanatomía funcional.
Los ERP son ampliamente interpretados como la suma de actividad cerebral causada por estimulación sensorial (Cruikshank et al., 2010; Kok, 1997; Luck, 2014; Muluh, 2011). En la Figura 1 se implementa la Ecuación 1 utilizando distintos valores de n y permite apreciar que el ERP no necesariamente refleja la complejidad de los diferentes ensayos que lo componen. Es decir, es un pobre descriptor de las dinámicas de potenciales postsinápticos excitatorios/inhibitorios de grandes ensambles neuronales (Makeig et al., 2004; Nunez & Srinivasan, 2006). A continuación, revisaremos distintos mecanismos propuestos que explican las fluctuaciones voltaicas de las que se deriva el ERP.

Figura 1. Ecuación 1 en distintos valores de n.
Esta figura muestra la gráfica de los cálculos realizados para distintos valores de n haciendo uso de la Ecuación 1.
ERP es la técnica más utilizada en electrofisiología cognitiva pero sus mecanismos fisiológicos se desconocen (Makeig et al., 2004; Makeig & Onton, 2009). Existen varios modelos generativos intentando explicar la emergencia de “componentes” ERP; fluctuaciones que presentan polaridad, amplitud, morfología, topología y latencias relativamente estables (Jervis et al., 1983).
Este modelo considera la actividad EEG en curso como ruido no correlacionado. Perturbaciones sensoriales o eventos cognitivos gatillan actividad neural. La nueva actividad se suma al ruido en curso, por lo que el promedio de múltiples ensayos aumenta la relación señal-ruido (signal-to-noise ratio), “rescatando” al ERP (Figura 2a).
Este modelo considera la actividad EEG en curso como actividad cerebral genuina con implicancias funcionales (He et al., 2010). Perturbaciones sensoriales o eventos cognitivos re-organizan transitoriamente esas señales (Figura 2b). Emergen nuevas estructuras funcionales, dependientes de poblaciones neuronales específicas, cuyo operar se manifiesta en fluctuaciones en el tiempo y la frecuencia (Brandt et al., 1991; Makeig et al., 2004). Esta nueva organización funcional da origen a los ERP al reiniciar o re-estructurar la fase del EEG en curso ensayo tras ensayo (Makeig et al., 2004; Makeig & Onton, 2009).
Tanto el modelo evocado como el de reinicio de fases son los que más aparecen en la literatura (Makeig et al., 2004; Min et al., 2007). Sin embargo, ninguno ha sido aceptado totalmente por la comunidad científica, resultando un continuo debate. De esta manera, se ha propuesto una solución parsimoniosa por medio de simulaciones computacionales: un modelo mixto entre la adición de poder y el reinicio de la fase en curso (Barry, 2009; Makeig et al., 2004; Makeig & Onton, 2009; Min et al., 2007). Así, los ERP se generarían tanto por efectos aditivos y no-lineales (David et al., 2006), como por cambios repentinos y transitorios en distintas bandas de frecuencia (Burgess, 2012).
Si bien la evidencia muestra que el modelo mixto sería una buena alternativa para explicar la emergencia del ERP (Min et al., 2007), aún existen datos que no pueden ser explicados. La literatura describe que la actividad EEG en curso (~10 Hz) tiene dinámicas de amplitud asimétricas (Mazaheri & Jensen, 2010) lo que implica que -algunas veces- el punto máximo de la oscilación se modula de forma diferente al punto mínimo de la misma señal, y viceversa (Figura 2c). Esta propiedad no-sinusoidal produce ondas lentas sin relación alguna con la integración funcional de estructuras cerebrales (Mazaheri & Jensen, 2010; Nikulin et al., 2010). Así, los ERP no darían cuenta de la formación de nuevas estructuras funcionales, si no que reflejan diferencias sistemáticas en la amplitud máxima que alcanza el EEG en curso en relación a su punto mínimo luego de la percepción de un estímulo (Mazaheri & Jensen, 2010). Debido a esto, una forma más adecuada de entender al EEG sería mediante su análisis ensayo a ensayo en el dominio del tiempo y la frecuencia.
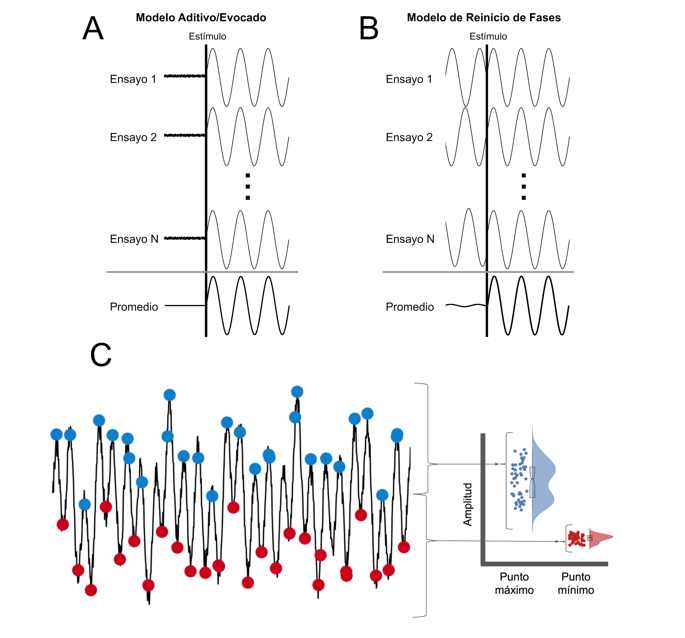
Figura 2a, b, c. Mecanismos generativos del ERP.
Esta figura grafica los resultados de tres mecanismos o maneras de realizar el cálculo de ERP.
Independientemente de los mecanismos generativos a la base de las dinámicas observadas en el EEG/ERP, la literatura describe una gran variedad de componentes ERP. Estos han sido consistentemente vinculados a funciones sensoriales, motoras y cognitivas (Bötzel & Grüsser, 1989; Donchin, 1979; Kok, 1997; Kutas & Federmeier, 2011; Luck, 2014; Näätänen & Michie, 1979; Picton, 1992; Pritchard, 1986; van Beijsterveldt & Boomsma, 1994). Por lo general, los componentes son denominados utilizando la letra “P” o “N” en mayúscula para indicar su polaridad -Positiva o Negativa- acompañados con un número que indica su latencia[8]. Entonces, un componente negativo cuya amplitud máxima respecto a la línea base ocurre alrededor de los 170 ms post-estímulo, se le denominará “N170”. Si bien múltiples componentes ERP han sido descritos a lo largo de la historia (Hillyard & Kutas, 1983; Luck, 2014), a continuación, describimos brevemente algunos de los más comunes. Además, mostramos el estándar para reportar ERP en la literatura actual (Figura 3).
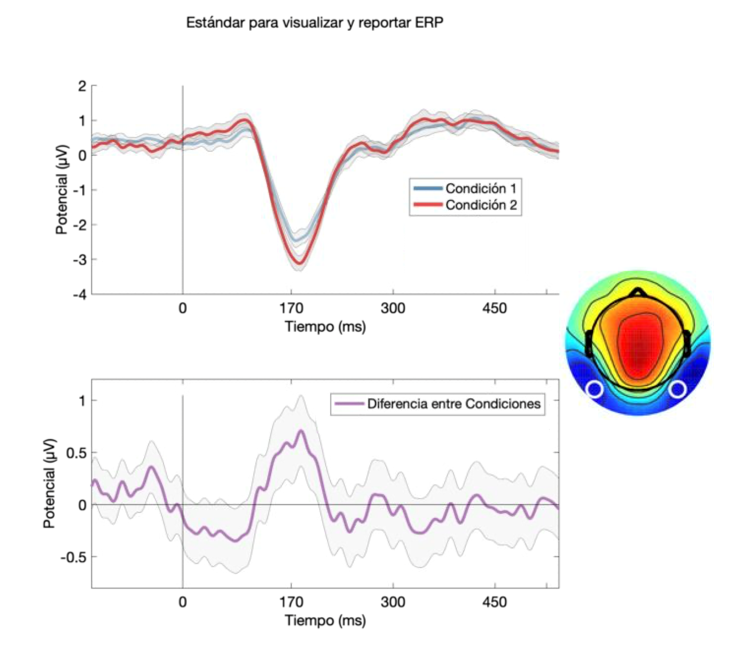
Figura 3. Estándar para visualizar y reportar ERP.
Representación estándar de la forma de visualizar y reportar ERP.
Topográficamente localizado en electrodos posteriores, se caracteriza como una pequeña deflexión sensible a propiedades físicas de los estímulos (e.g. contraste, frecuencia espacial). Temporalmente extendido entre 40-60 y 120-140 ms post-estimulación. Su amplitud máxima es alcanzada alrededor de 80-100 ms y su polaridad varía pues sería generado en la corteza visual primaria (V1) plegada en la cisura calcarina, de modo que si es registrado sobre ella será positivo (estímulo en el campo visual inferior) y viceversa (Martı́nez et al., 2001).
Topográficamente localizado en electrodos latero-occipitales, depende tanto de las propiedades físicas del estímulo como del nivel de arousal del participante. Temporalmente extendido entre 60-90 y 130-150 ms post-estimulación. Con latencia positiva, su amplitud máxima se alcanza alrededor de 80-120 ms. Su origen cortical se ha estimado en el giro occipital medial de la corteza extraestriada con extensiones hacia áreas ventrales como el giro fusiforme (Luck, 2014; Martı́nez et al., 2001). La versión auditiva del P1/P100 presenta amplitud máxima cerca de los 50 ms en electrodos frontocentrales (Luck, 2014).
Topográficamente localizado en la región medial de la corteza frontal, refiere a una activación que se presenta posteriormente al cometer un error (Gehring et al., 1993; Phan Luu et al., 2003). La corteza cingulada anterior se ha visto involucrada en la generación de una onda de actividad cerebral eléctrica negativa (Falkenstein et al., 1991; Gehring, 1993; Herrmann et al., 2004; Phan Luu et al., 2003) que se presenta aproximadamente 100 ms tras cometer un error (Falkenstein et al., 1991; Gehring et al., 1993) con amplitud de aproximadamente 10 μV (Dehaene et al., 1994; Stemmer et al., 2004). Esta actividad cerebral se ha observado en tareas de discriminación/elección y tiempo de reacción al cometer errores (errores de elección, de acción y de no-acción) (Luu et al., 2000; Scheffers et al., 1996), en comparación con la actividad cerebral al entregar respuestas correctas (Falkenstein et al., 1991; Gehring et al., 1990, 1993; Stemmer et al., 2004).
Son manifestaciones opuestas del mismo dipolo, topográficamente localizados en sensores occipitales (N1/N170) y en la línea media central (VPP). Temporalmente extendidos entre 100-200 ms post-estimulación, alcanza amplitud máxima a los 170 ms con polaridad negativa (N170) y positiva (VPP). El N170 es una instancia especial del N1, específico para rostros (Bötzel & Grüsser, 1989). La atención espacial influye sobre estos componentes tanto a nivel visual como auditivo y han sido localizados en la fisura calcarina (Luck, 2014).
Topográficamente localizado en sensores parieto-centrales y de polaridad positiva, depende de estímulos simples que han sido previamente identificados como objetivos perceptuales y/o conductuales de carácter infrecuente (Luck, 2014). Temporalmente se extiende después de 200 ms post-estimulación.
Topográficamente localizado en sensores de la línea media central, depende de la interrupción de una regla establecida por una secuencia de estímulos sensoriales independiente de la atención (Näätänen, 2001). Temporalmente extendido en 160-220 ms post-interrupción.
Topográficamente localizado en sensores fronto-parietales, depende de la detección de errores conductuales, identificación de estímulos infrecuentes que representen metas conductuales y procesos cognitivos complejos sensibles al contexto de la tarea. Temporalmente extendido entre 200 y 500 ms, no tiene amplitud máxima dada su morfología extensa de polaridad positiva (Karlin et al., 1971; Luck, 2014).
Topográficamente distribuido sobre electrodos parieto-occipitales, depende del procesamiento de significado semántico lingüístico y no-lingüístico, lenguaje y memoria semántica (Kutas & Federmeier, 2011; Kutas & Hillyard, 1980). Con polaridad negativa, se extiende temporalmente entre 250 y 500 ms post-estimulación.
Topográficamente localizado en electrodos centro-parietales, se conoce también como potencial pre-motor o de preparación (readiness). Se asocia a la ejecución de actos motores deliberados (Kornhuber & Deecke, 1965, 1964). En contraste con los otros potenciales descritos, BP se calcula alineando las señales al momento de comienzo del acto motor de la persona -típicamente, mover un dedo de la mano- en intervalos irregulares, de forma deliberada y voluntaria. El BP temprano comienza ~3 s antes del inicio del movimiento, mientras que el BP tardío comienza ~500 ms. Este intervalo de tiempo (entre la detección de un evento eléctrico y el estar consciente de su decisión), ha generado discusiones en torno a la existencia del libre albedrío (Libet, 1985).
En la literatura académica, varios de los componentes ERP han sido evaluados en distintas poblaciones clínicas, rangos etarios y con propósitos diferentes (diagnósticos, evaluación clínica, monitoreo de evolución, entre otros). A continuación, se presentan algunas de las implicancias y usos clínicos de componentes ERP.
El componente P100 se ha estudiado con relación a la ansiedad social y el movimiento ocular (Ran & Chen, 2017; Schmitz et al., 2012), evidenciando que los participantes que presentaban un alto nivel de ansiedad social presentan amplitudes más altas en P100, relacionadas con la mirada evitativa/desviada que aquellos participantes con bajo nivel de ansiedad social. Además, evidenciaron menores latencias tardías, lo cual sería un indicador de sesgo de procesamiento específico para la mirada desviada. Asimismo, Keil et al. (2018) estudiaron el componente N170 en niños y adolescentes respecto a procesos de control cognitivo en tareas de reconocimiento facial, encontrando que en los niños y adolescentes que presentaban Trastorno de Ansiedad Social, el componente N170 tenía amplitud menor respecto al reconocimiento de caras neutrales.
Por otro lado, el componente Mismatch Negativity (MMN) ha sido estudiado en distintas poblaciones y ámbitos clínicos, siendo utilizado principalmente para monitorear la evolución de pacientes comatosos y predecir su despertar (Duncan et al., 2009). MMN también se ha estudiado en pacientes que presentan diagnóstico de Esquizofrenia (Duncan et al., 2009; Michie, 2001), evidenciando una disminución de la amplitud de MMN en comparación con pacientes con diagnóstico de bipolaridad y controles sanos (Salisbury et al., 2007). Este componente también ha sido utilizado como índice de deterioro cognitivo en patologías como Esclerosis Múltiple y pacientes que presentan afasia (Duncan et al., 2009; Ilvonen et al., 2003; Jung et al., 2006).
P300 ha sido estudiado en pacientes con demencia para la evaluación del funcionamiento cognitivo, encontrando que el peak de latencia de este componente en pacientes con demencia era más prolongado en comparación con controles sanos (Duncan et al., 2009; Goodin et al., 1978). El componente P300 también ha sido utilizado en investigaciones vinculadas que tiene como objetivo estudiar recursos atencionales, trastornos anímicos, dependencia al alcohol, demencias y sobrevivientes de lesiones cerebrales traumáticas y otro tipo de diagnósticos presentes en poblaciones jóvenes; a saber: Espectro Autista y Trastorno por Déficit Atencional e Hiperactividad (Duncan et al., 2009).
Respecto al uso en evaluación clínica del componente N400, cabe destacar que tanto los decrementos en su amplitud y las demoras de latencia se han estudiado respecto a la presencial y severidad de déficits que involucran la comprensión (Connolly & D’Arcy, 2000; Friederici et al., 1998; Marchand et al., 2002). Además, el componente N400 ha sido estudiado con relación a los efectos del contexto semántico en pacientes que presentan Alzheimer, así como también para el estudio de síntomas significativos de la esquizofrenia como lo es el discurso desorganizado (Duncan et al., 2009).
Asimismo, el componente Error-Related Negativity (ERN) se ha asociado a diferencias individuales en la impulsividad del estilo de respuesta y la angustia (Gehring et al., 2000; P. Luu et al., 2000; Pailing et al., 2002; Stemmer et al., 2004). ERN se ha estudiado en niños con Trastorno Obsesivo-Compulsivo y Trastorno de Ansiedad Generalizada, evidenciándose un aumento del componente ERN en comparación con controles (Gehring et al., 2000; Hajcak et al., 2008; Ladouceur et al., 2006; Olvet & Hajcak, 2008). Además, se mostró que la amplitud de ERN no presentó cambios significativos luego de la realización de tratamientos exitosos para Trastornos Obsesivos-Compulsivos (Hajcak et al., 2008), indicando que este componente no estaría determinado por cambios en la sintomatología ansiosa (Olvet & Hajcak, 2008). Otras poblaciones clínicas también han sido estudiadas respecto al componente ERN, como por ejemplo con pacientes que presentan Depresión (Elliott et al., 1998; Holmes & Pizzagalli, 2007; Olvet & Hajcak, 2008; Pizzagalli et al., 2006), Esquizofrenia (Henderson et al., 2006; Mathalon et al., 2002) y niños con diagnóstico del Espectro Autista (Henderson et al., 2006; McPartland et al., 2004; Townsend et al., 2001).
Finalmente, el BP -junto con toda la familia de potenciales corticales relacionados al movimiento (Shakeel et al., 2015)- han sido estudiados en contextos clínicos. La evidencia sugiere que modulaciones en la amplitud del BP se vincula a lesiones en el tracto piramidal vinculado a la corteza motora primaria (Kitamura et al., 1993) y el giro supramarginal en el lóbulo parietal y sus proyecciones al lóbulo frontal (Wheaton et al., 2005). Además, en el contexto de sobrevivientes de Accidentes Cerebrovasculares, la amplitud de BP se asocia a la recuperación post-accidente (Gerloff et al., 2006). Asimismo, utilizando un paradigma de ejecución de movimientos sincronizados, BP se manifiesta bilateralmente; indicando una activación de las cortezas motoras bilaterales (Mayer et al., 1995). Cuando los movimientos son ejecutados unilateralmente y BP se expande bilateralmente, puede ser tomado como un indicador de pérdida de inhibición transcallosa (Maegaki et al., 2002).
A principios del siglo XX, la medición de la actividad eléctrica del cerebro a gran escala emerge como un objeto epistemológico para la Psicología y la Neurociencia. Comenzando, así, la Electrofisiología Cognitiva como subdisciplina que busca describir y entender los correlatos electrofisiológicos de la percepción, la acción y los procesos cognitivos. La electrofisiología cognitiva continúa su desarrollo hoy en día -con importantes desafíos y oportunidades- al aplicar nuevas técnicas de análisis temporales, espectrales y topográficos. Este contexto permitirá mayor entendimiento sobre los mecanismos subyacentes a la emergencia de los patrones de actividad eléctrica cerebral, que sistemáticamente reflejan procesos perceptuales, cognitivos y de interacción con el medio.
Los autores agradecen a J.I. Egaña, Ph.D. por sus relevantes comentarios sobre el manuscrito original. Asimismo nos gustaría agradecer a D. Rojas-Líbano, Ph.D. por su contribución en el párrafo de Bereitschaftspotential (BP) y sus comentarios que mejoraron las figuras del presente manuscrito.
Contribuciones específicas de los autores: A.G. C. llevó a cabo revisión bibliográfica, editó y escribió el manuscrito final. J.A. llevó a cabo la revisión bibliográfica inicial en el contexto de la realización de su tesis para optar al grado de Magíster en Neurociencia Social y editó versiones tempranas del manuscrito. M.A.F.G. apoyó y complementó la revisión bibliográfica y editó ediciones tempranas del manuscrito. F.J.P. supervisó la revisión bibliográfica, escribió y editó el manuscrito final, hizo las figuras y escribió la función de MATLAB que se adjunta en como material complementario.
El presente trabajo fue financiado por el proyecto Nº11180620 del concurso Iniciación en Investigación del Fondo Nacional de Desarrollo Científico y Tecnológico (FONDECYT), de la Agencia Nacional de Investigación y Desarrollo (ANID) del Ministerio de Ciencia, Tecnología, Conocimiento e Innovación del Estado de Chile.
Conflictos de interés:Los autores declaran no tener conflictos de interés.
function [ERP]=CalculaERP(datosCrudos,vectorTiempo,hacerGraficos)
% Nombre del programa: CalculaERP.m
% Uso: [ERP]=CalculaERP(datosCrudos,vectorTiempo,hacerGraficos);
%
% Variables de entrada:
% datosCrudos. Es una variable (vector o matriz) que
% contiene al menos 1 sensor de datos de EEG y sus respectivos ensayos.
% El orden de este vector o matriz sigue la nomenclatura del formato EEGLAB
% (Delorme, A., & Makeig, S. (2004). EEGLAB: an open source toolbox for
% analysis of single-trial EEG dynamics including independent component
% analysis. Journal of neuroscience methods, 134(1), 9-21.) donde los datos
% se guardan en un vector/matriz de 3 dimensiones donde la primera dimensi?n
% corresponde a sensores, la segunda es puntos de datos (tiempo) y la
% tercera corresponde a ensayos experimental.
%
% vectorTiempo. Vector que contiene el curso temporal del segmento de datos
% que se est? calculando. Esta variable es opcional.
%
% hacerGraficos. Es un l?gico (0|1) para indicar si el usuario desea
% graficar el ERP calculado. Al ingresar 0, no se realizar?n gr?ficos, al
% ingresar 1, un gr?fico parecido al panel superior de la Figura 3 (l?nea
% 293) aparecer?. Si los datos ingresados contienen m?s de 1 sensor, la
% figura ser? un "butterfly plot" (todos los sensores sobrepuestos en una
% misma figura.
%
% Variable de salida:
% ERP. Es una variable (vector o matriz) que contiene
% el mismo n?mero de sensores que la variable datosCrudos. Su orden sigue
% la nomenclatura EEGLAB. En esta oportunidad el vector/matriz tendr? una
% dimensi?n menos que la variable datosCrudos, ya que la dimensi?n de
% ensayos experimentales habr? sido promediada para computar el ERP. Esta
% variable puede ser graficada o testeada estad?sticamente utilizando
% MATLAB o alg?n otro software estad?stico.
%
% Al ingresar el vector/matriz con datos, la funci?n proceder? a computar
% el ERP al implementar la Ecuaci?n 1 del art?culo (l?nea 160).
%
% Nota importante: Para ejecutar esta funci?n computacional se requiere
% que el ordenador tenga instalado el ambiente MATLAB
% (The MathWorks, Inc., Natick, MA).
% Funci?n escrita en Abril de 2019 por Francisco J. Parada, Ph.D.
% (francisco.parada@udp.cl) de la Facultad de Psicolog?a de la Universidad
% Diego Portales para su difusi?n como documento suplementario del art?culo
% "Computaci?n, an?lisis e interpretaci?n de se?ales el?ctricas del cerebro
% humano en el dominio del tiempo: 90 a?os de Electrofisiolog?a Cognitiva".
% Junto con la presente funci?n computacional, se provee el archivo
% datosCrudos.mat que contiene 11 sensores occipitales junto con 748 puntos
% de datos de EEG en 101 ensayos experimentales. Estos datos fueron
% recolectados en el sistema Electrical Geodesics Inc. (EGI) que se
% encuentra ubicado en el Laboratorio de Neurociencia Cognitiva y Social de
% la Facultad de Psicolog?a de la Universidad Diego Portales, en Vergara
% 275, Santiago de Chile.
%% Calcula ERP
close all; %cierra todas las figuras anteriores
if isempty(datosCrudos) %comprueba si datosCrudos contiene datos
error('datosCrudos parece no contener datos. Revisar y volver a intentar.')
return
end
if size(datosCrudos,3)>1 %comprueba la existencia de dimensiones
ERP=mean(datosCrudos,3); %Promediar la tercera dimensi?n de ensayos experimentales
elseif size(datosCrudos,2)>1 %comprueba las dimensiones
ERP=mean(datosCrudos,2); %Promediar la tercera dimensi?n de ensayos experimentales
end
if ~exist('vectorTiempo','var') %comprueba si el par?metro opcional fue ingresado
vectorTiempo=1:length(ERP);
end
if hacerGraficos %comprueba si se graficar? el resultado
fig=figure; %abre una nueva figura
fontSize=15; %establece tama?o de fuente
plot(vectorTiempo,ERP','linewidth',3);
xlabel('Tiempo (en milisegundos)','FontSize',fontSize);
ylabel('Potencial (en microvolts)','FontSize',fontSize);
title('Potencial relacionado a eventos (ERP)')
end
end
francisco.parada@udp.cl
Figura 1. Ecuación 1 en distintos valores de n.
Esta figura muestra la gráfica de los cálculos realizados para distintos valores de n haciendo uso de la Ecuación 1.
Figura 2a, b, c. Mecanismos generativos del ERP.
Esta figura grafica los resultados de tres mecanismos o maneras de realizar el cálculo de ERP.
Figura 3. Estándar para visualizar y reportar ERP.
Representación estándar de la forma de visualizar y reportar ERP.


