Carátula del artículo
Estudio comparativo de la comunidad de
moluscos y crustáceos epifaunales en los manglares La Honda y Che Paulito, Los Santos, Panamá, 2019 – 2020
Comparative study of the epifaunal molluscs and
crustaceans community in La Honda and Che Paulito mangroves, Los Santos, Panamá, 2019-2020
 Virgilio Villalaz virgilio.05vd@gmail.com
Virgilio Villalaz virgilio.05vd@gmail.com
Universidad de Panamá, Panama
Nelva Villarreal nelva.villarreal@up.ac.pa
Universidad de Panamá, Panama
Juan Antonio Gómez H. juanay05@hotmail.com
Universidad de Panamá, Panama
Italo Goti italo.goti@up.ac.pa
Universidad de Panamá, Panama
Visión Antataura
Universidad de Panamá, Panamá
ISSN: 2309-6373
ISSN-e: 2520-9892
Periodicidad: Semestral
vol. 6, núm. 2, 2022
Recepción: 07 Agosto 2022
Aprobación: 09 Noviembre 2022
1. Introducción
El ecosistema de manglar es un ambiente altamente
productivo, basado en el aporte de
hojarasca, el cual alcanza 712 g/m2/año en manglares áridos de
México (Sánchez-Andrés et al., 2010) hasta 12 629 kg/ha/año en manglares ribereños en
Panamá (Rodríguez et al., 2012). Este nivel de productividad proveniente de las hojas, así como el aporte de fitodetrito de los tributarios (Bouillon et al., 2002) es la base para el establecimiento de una alta diversidad de invertebrados en este ecosistema, que a pesar de estar representados por diversos grupos como: Echinodermata, Cnidaria, Porífera, Polychaeta, Molusca y Crustáceos (Quiceno Cuartas y Palacio Baena, 2008), los Arthropoda y Molusca son los grupos más diversos, según García Padilla y Palacio (2008). Una de las características del sitio de estudio es el bajo desarrollo del bosque de manglar por su localización en el área del arco seco, aledaño a una finca de acuicultura, que de acuerdo a la presencia de muros de estanques camaroneros, producen un efecto de borde sobre la diversidad y abundancia de los invertebrados. Este efecto antropogénico induce a respuestas complejas en las comunidades de invertebrados debido a la ruptura de la estructura del bosque de manglar (Berguer et al., 2008). Sin embargo, Sartizabal et al., (2012) indican que, en áreas intervenidas, a pesar de que la diversidad no varía con respecto a manglares prístinos, la composición (Error 1: Reference must be linked to an existing reference) (Error 2: Type of reference is a required field) (Error 3: Related URL doesn't exist)
de especies, la equitatividad y dominancia sí se ven afectadas.
Las comunidades de invertebrados del ecosistema de manglar presentan variaciones en la densidad y abundancia de las especies entre zonas, dentro del ecosistema, y entre temporadas del año, lo que promueve la dominancia debido a la presencia de un número reducido de especies con capacidad adaptativa a las variaciones del medio (Jiménez, 1994). Se diferencia de la comunidad de playas arenosas por estar sujeta a factores como: disponibilidad de nutrientes, reclutamiento larvario, características ambientales como la desecación y variación térmica (Pfaff y Nel, 2019). Sin embargo, tanto la abundancia como la diversidad también pueden estar sujetas al estrés producido en ambientes fragmentados o contaminados (Cárdenas-Calle y Mair 2014).
En el caso de los manglares La Honda y Che Paulito, se desconoce la estructura de la comunidad de moluscos y crustáceos y las variaciones interanuales, razón que explica el interés de estudiar la dinámica de estos invertebrados en dichos manglares, con el propósito de descifrar estos componentes comunitarios durante los años 2019 y 2020.
2. Materiales y métodos
El estudio se llevó a cabo en los manglares: La Honda
ubicado en la siguiente coordenada (N572674,85 m E8751011,13 m) y Che Paulito (N 572321,52 m E 876104,25 E), a una distancia de 1,39 km entre ellos y a 3,17 km y 3,51
km de la comunidad de Santa Ana de Los Santos, respectivamente, en los cuales se establecieron dos cuadrantes de 100 m2 cada uno, por manglar (figura 1).
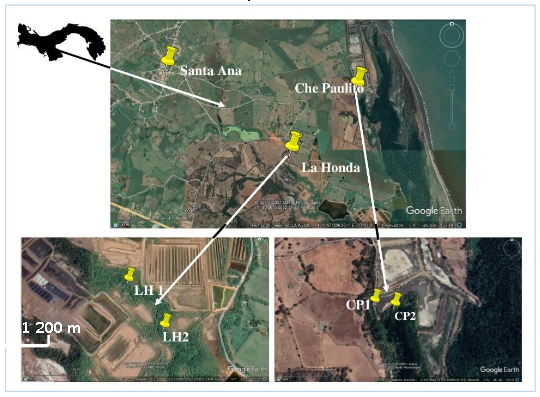 Figura 1.
Mapa de localización de
la zona de estudio, manglares La Honda y Che Paulito, Los Santos.
Google Earth, modificada por los
autores. Imagen NES/Airbus.https://www.nativatours.com/es/los-santos-3/
Figura 1.
Mapa de localización de
la zona de estudio, manglares La Honda y Che Paulito, Los Santos.
Google Earth, modificada por los
autores. Imagen NES/Airbus.https://www.nativatours.com/es/los-santos-3/
Las muestras de moluscos y crustáceos de la epifauna, de dos cuadrantes por manglar, se tomaron a mano libre, en un muestreo mensual, durante los meses de junio a agosto, en dos campañas: la primera en 2019 y la segunda en 2020, en períodos de marea baja de sicigia de luna llena. Los organismos colectados fueron fotografiados para su identificación y devueltos al sustrato para minimizar el impacto sobre el ecosistema.
La abundancia se expresó en términos de número de individuos/600 m2, producto de la suma de dos cuadrantes mensuales de todos los organismos colectados durante el período de muestreo, por manglar. Los organismos se identificaron en campo con la ayuda de las claves: Olson (1961), D’Croz (1992), Jiménez (1994), Fischer et al. (1995), Orellana Amador (2010) y Ross et al. (2014). (Error 4: Olson (1961) is linked to an element that doesn't exist)
Se calculó diferencias de abundancia entre períodos de colecta mediante el análisis U de Mann-Whitney, previa evaluación de la normalidad y de la proporción de cada taxón con la prueba de comparación de proporciones de muestras independientes (Steel y Torrie, 1985); la estructura de la comunidad se determinó mediante histograma de frecuencia del conteo de organismos por taxón, para definir la dominancia de los principales grupos.
La diversidad se estimó con el índice de Shannon-Wiener, basado en el logaritmo de base 10, se utilizó la prueba t de Hutcheson para determinar diferencias de diversidad, (Moreno, 2001). Se construyó la curva de rarefacción según Sanders (1968) y la rutina ANOSIM según Clarke y Warwick (2001), análisis utilizados para determinar la diferencia entre años; además se investigó la equitatividad de Pielou y los índices de Jaccard y Sorensen cualitativo para evaluar la similaridad (Gómez Herrera et al., 2022). Los datos fueron analizados con los paquetes libres BioEstat 3.1 (Ayres et al., 2007) y PAST 4.0 (Hammer et al., 2001).
3. Resultados Manglar La Honda
Se recolectaron en total 5 314 individuos durante los dos períodos, de los cuales 2 638 en el año 2019, representados por 12 taxones, de estos tres fueron gastrópodos (3,52 % de la abundancia) y nueve crustáceos (96,47 %). En el 2020 se obtuvieron 2 676 organismos, representados por 11 taxones, tres resultaron ser moluscos (3,55 %) y ocho crustáceos (96,44 % de la abundancia).
La especie dominante en el año 2019 fue Uca sp1, representó el 84,42 % de la abundancia, mientras que las abundantes estuvieron representadas por: Goniopsis cruentata, Hemigrapsus sp, Uca sp2, Cerithidiopsis montagnei y C. californica (figura 2).
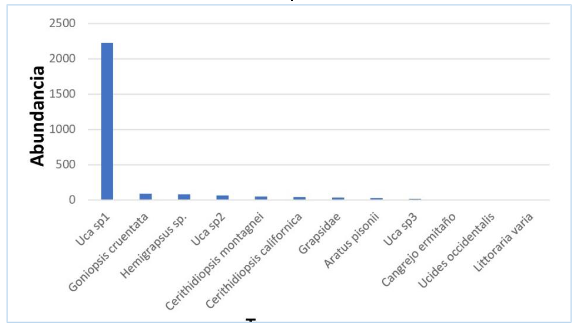 Figura 2.
Estructura
de la comunidad de crustáceos y moluscos en el manglar La Honda, 2019
Los autores.
Figura 2.
Estructura
de la comunidad de crustáceos y moluscos en el manglar La Honda, 2019
Los autores.
En 2020, la especie dominante fue Uca sp1, que representó el 84,41 % de la abundancia, mientras que entre las especies abundantes tenemos Goniopsis cruentata, Uca sp2, Grapsidae,
Cerithidiopsis montagnei y C. californica (figura 3).
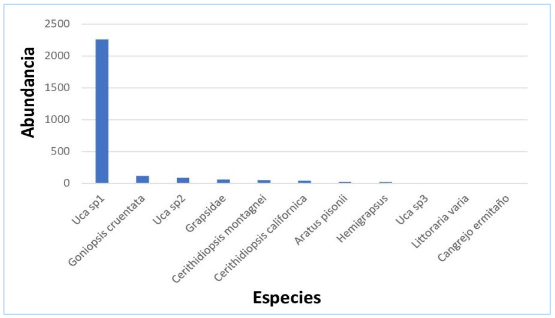 Figura 3.
Estructura de la
comunidad de crustáceos y moluscos en el manglar La Honda, 2020
Los autores
Figura 3.
Estructura de la
comunidad de crustáceos y moluscos en el manglar La Honda, 2020
Los autores
No se demostró diferencia de la abundancia (U = 63,5 p = 0,8777), ni de la proporción de ninguno de los taxones entre años (z < 1,96 p > 0,05).
El índice de diversidad de Shannon-Wiener indicó un valor de 2,0468 en el año 2019, mientras que en el 2020 fue de 1,9010, sin embargo, se obtuvo diferencia
altamente significativa entre estas (tH = 3,1548 p = 0,0016645). La equidad fue de 0,98 y 0,97 respectivamente para cada año. Las curvas de rarefacción mostraron mayor diversidad en 2019 (figura 4).
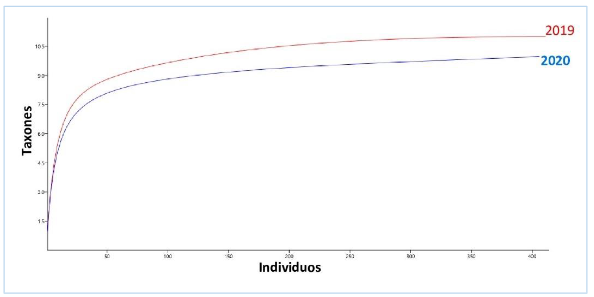 Figura 4.
Curva de rarefacción de moluscos y crustáceos del manglar La Honda en 2019 y 2020.
Los autores.
Figura 4.
Curva de rarefacción de moluscos y crustáceos del manglar La Honda en 2019 y 2020.
Los autores.
La prueba de ANOSIM marcó similaridad entre años (R =
-0,03362 p = 0,6604), el análisis multidimensional no métrico produjo un estrés de 0,05218, catalogado como un buen análisis (figura 5).
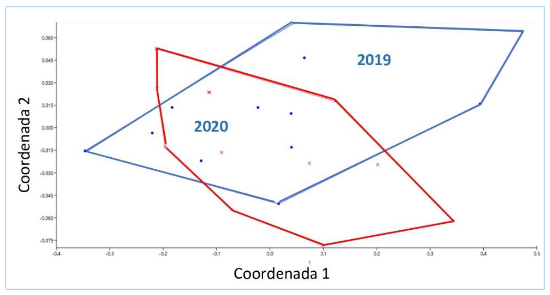 Figura
5.
Análisis multidimensional no métrico del manglar La Honda entre 2019 y 2020.
Los autores
Figura
5.
Análisis multidimensional no métrico del manglar La Honda entre 2019 y 2020.
Los autores
La similaridad fue de 0,92 según el índice de Jaccard y 0,96 de acuerdo al índice de Sorensen cualitativo.
Manglar Che Paulito
Se recolectaron 4 669 organismos en ambos períodos, 2 400 en el año 2019, representados por 22 taxones, de estos nueve resultaron ser moluscos, cuatro gastrópodos con el 7,71 % de la abundancia y cinco bivalvos (2,58 %), 13 crustáceos (89,71 % del total colectado). En el 2020 se obtuvieron 2 269 individuos, con 15 taxones, seis moluscos, de los cuales 3 gastrópodos representaron el 9,17 % de la abundancia y tres pelecípodos (1,81 %), además de nueve crustáceos (89,02 % del total de individuos). La especie dominante en 2019 fue Uca sp1, que representó el 75,04 % de la abundancia, mientras que entre las especies abundantes tenemos Cerithidiopsis californica. Goniopsis cruentata, Uca sp3, Hemigrapsus sp, Grapsidae, Anadara
tuberculosa y Aratus pisonii (figura 6).
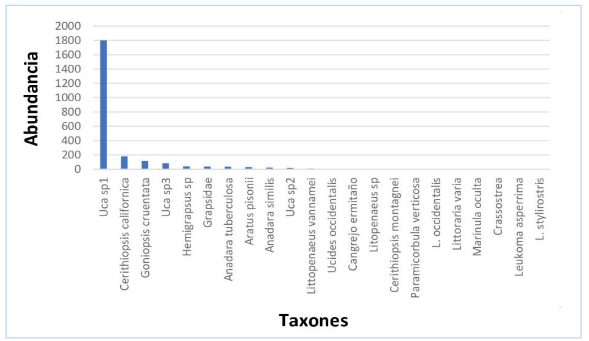 Figura 6.
Estructura de la comunidad de crustáceos y moluscos en
el manglar Che Paulito, 2019
Los autores
Figura 6.
Estructura de la comunidad de crustáceos y moluscos en
el manglar Che Paulito, 2019
Los autores
La especie dominante en 2020 fue Uca sp1, que representa el 69,99 % de la totalidad de individuos, mientras que entre las especies abundantes tenemos Cerithidiopsis californica. Goniopsis cruentata, Uca sp2, Grapsidae, Hemigrapsus sp, Uca sp3, Anadara tuberculosa y Aratus pisonii, que representaron el 27,06 %; el porcentaje restante, 2,95 %, correspondieron a las especies comunes y raras (figura 7)
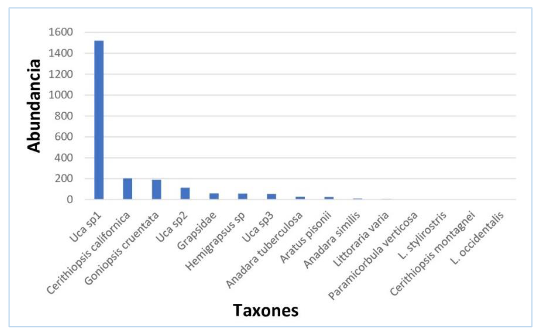 Figura 7.
Estructura de la comunidad de crustáceos y moluscos en
el manglar Che Paulito, 2020
Los autores
Figura 7.
Estructura de la comunidad de crustáceos y moluscos en
el manglar Che Paulito, 2020
Los autores
No se presentó diferencia significativa de la abundancia entre años en el manglar Che Paulito (U = 124,0 p = 0,2047), el único taxón que mostró diferencias de proporción en ese período fue Uca sp1 (z = 5,10 p < 0,05).
El índice de diversidad de Shannon-Wiener fue de 1,103 en el año 2019, mientras que en el 2020 fue de 1,2827, se obtuvo diferencia altamente significativa entre estos (tH = -4,296 p = 1,77 x 10-5). La equidad fue de 0,95 y 0,98 respectivamente para cada año. Las curvas de rarefacción mostraron mayor diversidad en 2019 (figura 8).
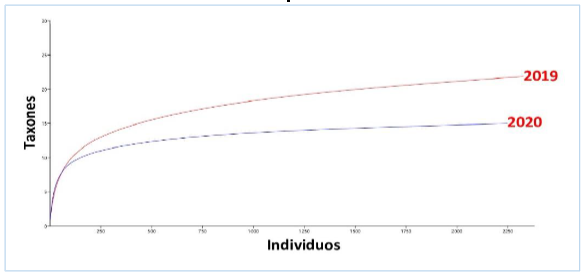 Figura 8.
Curva de rarefacción de moluscos y crustáceos del manglar Che Paulito en 2019 y 2020.
Los autores
Figura 8.
Curva de rarefacción de moluscos y crustáceos del manglar Che Paulito en 2019 y 2020.
Los autores
La prueba de ANOSIM mostró similaridad entre años (R = -0,005447
p = 0,4244), el análisis
multidimensional no métrico indicó un estrés de 0,08546, que se catalogó como buen análisis (figura 9).
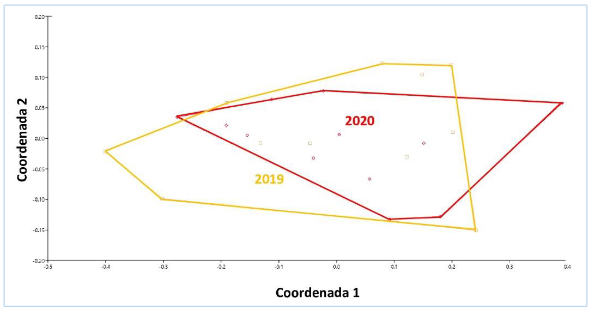 Figura 9.
Análisis
multidimensional no métrico del manglar Che Paulito, entre 2019 y 2020.
Los autores
Figura 9.
Análisis
multidimensional no métrico del manglar Che Paulito, entre 2019 y 2020.
Los autores
La similaridad fue de 0,68 según el índice de Jaccard y 0,81 de acuerdo al índice de Sorensen
cualitativo.
4. Discusión
Los ecosistemas de manglar proporcionan, por lo menos, 15 productos que garantizan la alimentación, vivienda y combustible a los moradores que viven en zonas aledañas a estos (Saint Paul, 2007), entre los grupos más frecuentemente encontrados en el manglar, que son útiles como fuente de proteína e importancia comercial, se encuentra variedades de crustáceos como los cangrejos, camarones y moluscos como Anadara tuberculosa, conocida como concha negra (Cornejo, 2014).
La abundancia de estos grupos de invertebrados, en los sitios estudiados, estuvo representada en su mayoría por crustáceos, y una proporción baja de moluscos, en contraste, en manglares ribereños de Indonesia Setyadi et al. (2021) reportaron 46 % de moluscos y 53 % de crustáceos; la proporción de este último taxón resultó ser inferior a lo encontrado en este estudio. No obstante, en la Bahía de Cispatá, Colombia, Quirós y Arias (2013), reportan dominancia de moluscos, con 85,2 % sobre crustáceos 14,8 %.
En particular entre los moluscos, en manglares de Asia es común encontrar, en la epifauna, una mayor abundancia de gastrópodos, al igual que en el presente estudio; en Indonesia, Baderan et al. (2019) reportan, en parcelas de 100 m2, 14 taxones, de los cuales 11 fueron gastrópodos, y 3 bivalvos, Rahardjanto et al. (2020), en parcelas de 500 m2, indican 11 gastrópodos y 2 bivalvos, Waran et al. (2020) en parcelas de 100 m2 sobre un transepto, en manglares multiespecíficos de Sonneratia sp, Rhizophora sp, Avicennia sp y Bruguera sp, recolectaron 9 gastrópodos y 2 bivalvos y Kaharundin y Wahidi (2020), en zonas dominadas por Bruguera sp, Rhizophora sp y Avicennia sp, obtuvieron 14 gastrópodos y 3 bivalvos. En Tailandia Premcharcoen et al. (2016) en manglares dominados por Avicennia sp y Rhizophora sp, identificaron los moluscos de la infauna mediante un nucleador, encontraron 12 gastrópodos y 6 bivalvos. Las proporciones arriba señaladas son semejantes a las reportada en esta investigación.
En el caso de los gastrópodos de la familia Littorinidae, estos predominan en áreas de regeneración joven de mangle (Macintosh et al., 2002), lo que explica la poca abundancia en los manglares de La Honda y Che Paulito, por ser un ecosistema muy intervenido, con características ecológicas de borde.
Por otro lado, la escasa abundancia de los bivalvos, se explica por la falta de movilidad, característica que facilita a los gastrópodos un amplio desplazamiento que les permite ocupar mayor espacio en el ecosistema (Kabir et al., 2014).
Macintosh et al. (2002) indican que la humedad retenida en el suelo es un factor que define la presencia de moluscos en este ecosistema. De acuerdo a Ashton et al. (2003) la abundancia de este grupo está definida por las características del subsuelo, como el potencial redox, el pH del líquido intersticial, sin embargo, entre los crustáceos, tanto la estructura de la comunidad de cangrejos, como la abundancia, está determinada por la altura topográfica del manglar.
Entre los crustáceos más abundantes en manglares de las costas del Pacífico Oriental Tropical, se encuentran los cangrejos del género Uca, conocidos como violinistas, comunes en zonas degradadas (Macintosh et al., 2002); la distribución de estos depende de la disponibilidad de factores físicos que determinan los nichos (Thurman et al., 2013). Entre los elementos que definen esta disposición tenemos: la
granulometría del sedimento, el contenido de materia orgánica y la retención de humedad en el suelo (Bezerra et al., 2006).
Las características antes mencionadas facilitan a los cangrejos Uca spp. la construcción de habitáculos para evadir la predación y refugiarse de variaciones ambientales, además de enterrar y macerar restos de hojas de mangle (Kristensen, 2008), lo cual incide sobre el potencial de oxi-reducción y percolación del líquido intersticial del sedimento (Penha-Lopes et al., 2009). El sistema de abrigo sirve como sumidero de metales traza y facilita su resuspensión, debido al enriquecimiento al estar disponible en mayor concentración en la capa superior de las madrigueras (Pan et al., 2022); además, mejoran la actividad aeróbica de las bacterias por la translocación de oxígeno (Mokhtari et al., 2016), por otro lado, Caparelli et al. (2022) señalan la importancia de los tubos de los cangrejos violinistas como fuente de depósitos de microplásticos en los procesos de bioperturbación del suelo, convirtiéndose en ingenieros del ecosistema, cuya actividad favorece la estructuración del manglar, aunado a los factores ambientales y geomorfológicos (Gupta y Verma, 2013).
Otros cangrejos, como la Familia Grapsidae, en manglares de Tailandia, dominan en zonas de bosque maduro, en el área de contacto con tierra firme (Kathiresan y Bingham, 2001), contrario a este estudio, donde la abundancia de esta familia es reducida por el poco desarrollo del manglar, debido a que han sido altamente intervenidos por la presencia de infraestructura de proyectos acuícolas. La importancia ecológica de los cangrejos de esta familia, considerados como uno de los taxones abundantes en la comunidad del ecosistema de manglar, se explica por el hecho de ser arborícolas (Fratini et al., 2005), estrategia utilizada por este taxón para disminuir la competencia, sin embargo, la escasa presencia en el sitio de estudio puede deberse al bajo desarrollo de esta vegetación.
Con relación a los cangrejos ermitaños, estos están sujetos a la disponibilidad, forma y tamaño de conchas, que en zonas tropicales no se constituye en un impedimento para la abundancia y distribución de dichos organismos (Teoh y Chong, 2014), sin embargo, De Grave y Barnes (2001) indican que su distribución depende de la complejidad del hábitat y la estrategia de predación de la cual son objeto.
La falta de complejidad del área y la escasez de conchas de gastrópodos como fuente de habitáculo para los cangrejos ermitaños, reportada en esta investigación, son razones que explican la poca abundancia de este taxón en la zona de estudio. El resto de los taxones resultó ser reducido posiblemente por la fragmentación, el efecto de borde y factores ambientales que pueden afectar a los manglares de La Honda y Che Paulito, reflejados en la diversidad encontrada.
Los valores del índice de diversidad calculado en ambos sitios revelan una alta biodiversidad, el índice de diversidad de Shannon-Wiener en el manglar de La Honda es mayor que los reportados, para la comunidad de moluscos y crustáceos, en Tailandia, con intervalo entre 0,03 y 1,36 (Macintosh et al., 2002) y en Indonesia, con valores de 0,62 a 1,19 (Setyadi et al., 2021). Por otro lado, para la comunidad de crustáceos, Márquez Rojas et al. (2006) comunican en rodales de Rhizophora spp de Venezuela un ámbito del índice entre 0,54 y 0,93, mientras que, en Brasil, Negromonte et al. (2012) indican variación entre 0,77 y 0,99.
En el manglar de Che Paulito el índice de diversidad presenta valores que están dentro del ámbito reportado por Satizabal et al. (2012) en el Pacífico colombiano, con amplitud entre 0,196 y 1,855, y del indicado para la comunidad de moluscos en Camerún, con valores entre 0,79 y 1,41 (Kottè-Mapoko et al., 2021), manglares cubiertos de las mismas especies de árboles que en Panamá. Hay que resaltar, que el índice de diversidad reportado para el manglar está por encima de los valores encontrados en la literatura, a pesar de las condiciones imperantes en el sector.
Con respecto al índice de equidad para ambos manglares, resultó ser elevado, lo cual no se explica debido a la dominancia de Uca sp1 en la comunidad. El índice de Pielou es superior a los valores reportados por Macintosh et al. (2002) en Tailandia, entre 0,13 y 0,76 y en Indonesia, donde varió de 0,47 a 0,92 (Setyadi et al., 2021). Kottè-Mapoko et al. (2021), en moluscos en Camerún, señalan vales de 0,66 a 0,91. Para la comunidad de crustáceos en Venezuela, Márquez Rojas et al. (2006) reportan un intervalo entre 0,66 y 0,94. Sin embargo, los valores aquí reportados están dentro del ámbito indicado por Satizabal et al. (2012) en Colombia, de 0,25 a 0,98 y Negromonte et al. (2012) en Brasil, con intervalo entre 0,89 a 0,95.
El efecto de borde antropogénico, debido a la construcción de estanques camaroneros, cercanos a las parcelas de estudio, ha facilitado la pérdida de cobertura de manglar y la consecuente degradación del ecosistema, tal como lo señala Arroyave- Rincón et al. (2014). La escasa cantidad de gastrópodos encontrados se debe precisamente a la degradación de los nichos a través de los cambios de la estructura del sustrato (Blanco y Castaño, 2012), que pueden incidir en los organismos debido a los requerimientos ambientales en el manglar. Cannicci et al. (2008) señalan que este grupo es sensible a los cambios producidos por las modificaciones permanentes sufridas al borde de este ecosistema. Igual sucede con los Crustáceos, como lo indica Arroyave- Rincón et al. (2014) en manglares del Golfo de Urabá, Colombia.
Según Amortegui-Torres et al. (2003) el efecto de borde expone a los organismos a temperaturas más altas, alteraciones en el sustrato y menor complejidad del hábitat, lo que conduce a una menor biodiversidad, sin embargo, en este caso, la consecuencia observada fue sobre la dominancia y no en la equidad y la diversidad.
A pesar del efecto de borde, la diversidad resultó ser alta. Es probable que los valores aquí obtenidos se atribuyan al método de muestreo sobre el suelo y a la forma de escoger los organismos; es posible que la técnica utilizada en la captura facilite la recolecta de los mismos, lo cual puede ir en detrimento de aquellos que están enterrados en el sustrato.
La diferencia de la diversidad encontrada entre los períodos de colecta (2019 – 2020) en los manglares de La Honda y Che Paulito, no se le puede atribuir al efecto del aumento en la presión de captura de invertebrados en los manglares por parte de los moradores de las comunidades costeras, que se vieron forzados a buscar recursos alimentarios por causa de las restricciones emanadas por el estado, debido al confinamiento producto de la pandemia por el SARCOV-2, indicadas por el Decreto Ejecutivo 490 de marzo de 2020 (Gaceta Oficial, 2020), que estableció el toque de queda en todo el país, por lo cual se esperaría un efecto sobre la abundancia y diversidad de moluscos y crustáceos en el manglar.
Esta diferencia, tampoco puede ser atribuida a fenómenos naturales como El Niño y La Niña, toda vez que el índice ONI de la NOAA, mostró en ambos periodos de colecta, características típicas de ENOS neutral, a excepción del trimestre mayo-junio-julio 2019,
Import Image con valores que definen el fenómeno ENOS cálido (El Niño), mientras que el trimestre julio-agosto-septiembre 2020 marcó ENOS frío (Las Niña), de acuerdo a la base de datos del Climate Prediction Center de la NOAA (https://origin.cpc.ncep.noaa.gov/products/analysis_monitoring/ensostuff/ONI_v5.pp) Fenómenos climáticos como el ENOS Cálido (El Niño) y ENOS Fríos (La Niña), son responsables de variaciones interanuales importantes en los ecosistemas marinos, sobre todo en los manglares, ya que afectan directamente la concentración de la salinidad del agua intersticial, por la escasez o abundancia de precipitación (Bolívar-
Anillo et al., 2020).
5. Conclusiones
· En ambos manglares el taxón más abundante fue los crustáceos. Entre los moluscos, los gastrópodos resultaron ser más variados, pero poco abundantes.
· La diversidad de los taxones estudiados fue más elevada que en manglares de la región
· El estudio comparativo de los dos períodos mostró igual abundancia, pero mayor diversidad durante el año 2019; sin embargo, las variaciones interanuales de la diversidad no pueden adjudicarse a los fenómenos naturales como El Niño/La Niña, ni al esfuerzo de captura por parte de la población aledaña debido al confinamiento producto del SARCOV-2.
Referencias bibliográficas
Ashton, E.C., Macinthosh, D.J. y Hogarth, P.J. (2003). A baseline study of the diversity and community ecology of crab and mollusks macrofauna in the Sematan mangrove forest, Sarawak, Malaysia. J. Trop. Ecol., 19(2), 127-142. https://doi.org/10.1017/S0266467403003158
Amortegui-Torres, V., Taborda-Marín, A.T. y Blanco, J.F. (2013). Edge effect on a Neritina virginica (Neritimorpha, Neritinidae) population in a black mangrove stand (Magnoliopsida,
Avicenniaceae: Avicennia germinans)
in the Southern Caribbean. Pan-american J. Aquat. Sci., 8(2), 68-78.
http://panamjas.org/pdf_artigos/panamjas_8(2)_68-78.pdf
Arroyave-Rincón, A., Amortegui-Torres, V., Blanco-Libreros, J.F. y Taborda-Marín, A. 2014. Efecto de borde sobre la población del cangrejo azul Cardisoma guanhumi (Decapoda: Gecarcinidae) en el manglar de la bahía El Uno, golfo de Urabá (Colombia): una aproximación a su captura artesanal. Actual. Biol., 36(100), 47-57. http://www.scielo.org.co/scielo.php?script=sci_arttext&pid=S0304- 35842014000100006
Baderan, D.W., Hamidun,
Ramliutina, M.S., Rahim, S. y Dali, R. (2019). The abundance and diversity of mollusks in mangrove ecosystem at coastal area of North Sulawesi, Indonesia. Biodiversitas, 20(4), 987-993.
https://doi.org/10.13057/biodiv/d200408
Berger, U., Rivera-Monroy, V.H., Doyle, T.W., Dahdouh-Guebas, F., Duke, N.C., Fontalvo- Herazo, M.L., Hildenbrandt, H., Koedam,
N., Mehling U., Piou, C., y Twilley, R.R. (2008). Advances and limitations of individual-based models to analyze and predict dynamics of
mangrove forests: A review. Aquatic
Botany, 89(2), 260-274. https://doi.org/10.1016/j.aquabot.2007.12.015
Bezerra, L.E.A., Dias, C.B.,
Santana, G.X. y Cascon, H.M. (2006). Spatial distribution of fiddler crabs (genus Uca) in a tropical
mangrove of northeast Brazil. Scientia Marina, 70(4), 759-766.
https://scientiamarina.revistas.csic.es/index.php/scientiamarina/article/view/82
Blanco, J.F. y Castaño, M.C. (2012). Efecto de la conversión del manglar a potrero
sobre
la
densidad y tallas de dos gastrópodos en el delta del río Turbo (golfo de Urabá, Caribe colombiano). Rev. Biol. Trop., 60(4), 1707-1719.
https://www.scielo.sa.cr/scielo.php?script=sci_arttext&pid=S0034- 77442012000400026
Bolivar-Anillo, H.J., Anfuso, G. y Abarca, S.C. (2020).
Eventos naturales y actuaciones antrópicas:
Impactos sobre el bosque de manglar de América del Sur. Revista Costas, 2(1), 211-232. https://doi.org/10.26359/costas.1802
Bouillon, S., Koedam, N., Raman, A.V. y Dehairs, F. (2002). Primary producers sustaining macro-invertebrate communities in
intertidal mangrove forests. Oecologia,
130, 441-448. https://doi.org/10.1007/s004420100814
Cannicci, S., Burrows, D., Fratini, S., Smith III, T.J.,
Offenberg, J. y Dahdouh-Guebas, F. (2008). Faunal impact on vegetation structure and ecosystem function in mangrove forests: A review. Aquat. Bot., 89, 186-200. https://doi.org/10.1016/j.aquabot.2008.01.009
Caparrelli, M.V., Martínez-Colón, M., Lucas-Solís, O., Valencia-Castañeda, G., Celis- Hernández, O., Ávila, E. & Mouatlet,
G.M. (2022). Can the bioturbation activity of the fiddler crab Minuca rapax modify the distribution of microplastics in sediments? Mar. Poll. Bull. 180, 113798.
https://doi.org/10.1016/j.marpolbul.2022.113798
Cárdenas-Calle, M. y Mair, J. (2014). Caracterización de macroinvertebrados bentónicos de dos
ramales estuarinos afectados por la actividad industrial, estero Salado – Ecuador. Rev. Intrópica, 9, 118-128.
https://revistas.unimagdalena.edu.co/index.php/intropica/article/view/1439/81 9
Clarke, K.R. y Warwick, R.M. (2001). Change in marine communities. An approach to statistical analysis and interpretation. Plymouth Marine Laboratory. U.K. https://updates.primer-e.com/primer7/manuals/Methods_manual_v7.pdf
Cornejo, X. (2014). Árboles y arbustos de los manglares del Ecuador. FAO ONU-REDD, 48
p. https://biblio.flacsoandes.edu.ec/libros/digital/55818.pdf
D’Croz, L. (1992). Guía taxonómica y morfológica para los crustáceos decápodos panameños. Universidad de Panamá.
De Grave, S. y Barnes, K.A. (2001). Ecology of tropical hermit crabs (Crustacea Decapoda) a Quirimba
Island, Mozambique: a multivariate assemblage perspective. Trop. Zool., 14, 197-209. https://doi.org/10.1080/03946975.2001.10531152
Fischer, W., Krupp, F., Schneider, W., Sommer, C., Carpenter, K.E. and Niem, V.H. (1995). Guía FAO para la identificación de especies para los fines de pesca. FAO. https://decapoda.nhm.org/pdfs/38962/38962.pdf
Gaceta Oficial. (2020). Decreto Ejecutivo 490 del 17 de marzo de 2020. “Que declara toque de queda en Panamá y dicta otras disposiciones”. Gaceta Oficial. No. 28983-
A. Ministerio de Gobierno y Justicia, Panamá. https://www.gacetaoficial.gob.pa/pdfTemp/28983_A/77907.pdf
García Padilla, J.M. y Palacio, J. (2008). Macroinvertebrados asociados a las raíces sumergidas del mangle rojo (Rhizophora mangle). Gestión y Ambiente, 11(3), 55-
66. https://revistas.unal.edu.co/index.php/gestion/article/view/14020
Gómez Herrera, J.A., Liñero Arana, I., Villalaz
Guerra, J. y Goti, I. (2022). Manual de métodos y técnicas aplicados a la Ecología
y Biología Marina. Universidad de Panamá.
Gupta, V.K. y Verma, A.K. 2013. Fiddler crabs, bioturbation and ecosystem engineering. Animal diversity, Natural History and conservation. Chap. 2(9), 181-192. https://www.researchgate.net/publication/318013038_Fiddler_Crabs_Bioturbati on_and_Ecosystem_Engineering
Hammer, O., Harper, D.A.T. y Ryan, P.D. (2001). PAST: Paleontological statistics software package for
education and data analysis. Palaeontología
Electrónica, 4(1), 1-9. https://palaeo-electronica.org/2001_1/past/past.pdf
Jickling, N. (2017). Shrimp aquaculture in Aguadulce: Impacts on mangrove forest health and shrimp larvae populations in two
sites on the Salado coastline. Independent Study Project (ISP) Collection. 2729. STRI. https://digitalcollections.sit.edu/isp_collection/2729
Jiménez, J.A. (1994). Los manglares del Pacífico centroamericano. UNA, Costa Rica.
Kabir, M., Abodlfathi, M., Hajimoradloo, A., Zahedi, S.l., Kathiresan, K. y Goli, S. (2014). Effect of mangroves on distribution, diversity and abundance of mollusks in mangrove ecosystem: a review. AACL Bioflux, 7(4), 286-300. http://www.bioflux.com.ro/docs/2014.286-300.pdf
Kathiresan, K. y Bingham, B.L. (2001). Biology of mangroves and mangrove ecosystems.
Adv. Mar. Biol., 40, 81-251. https://doi.org/10.1016/S0065-2881(01)40003-4
Massou, V.M., Essomè-Koum, G.L., Nyamsi-Moussian, L.,
Konango-Samè,
Boumakari, A., y Din, N. (2021). Dynamic of mangrove associated mollusks in anthropized areas of the
Cameroon coastline. Open J. Ecol., 11,
565- 579. https://doi.org/10.4236/oje.2021.118036
Kristensen, E. (2008). Mangrove crabs as ecosystem engineers; with emphasis on sediment processes. J. Sea Res., 59, 30-43.
https://doi.org/10.1016/j.seares.2007.05.004
Macintosh, D.J., Ashton, E.C. y Havanon, S. (2002). Mangrove rehabilitation and intertidal biodiversity: a study in the Ranong mangrove ecosystem, Thailand. Est. Coast. Shelf Sci., 55, 331-345. https://doi.org/10.1006/ecss.2001.0896
Márquez Rojas, B., Blanco Rambla, J.P., Jiménez, M., Allen, T. (2006). Crustáceos asociados a las raíces del mangle rojo Rhizophora mangle (L.) en el Golfo de Santa Fe, Estado Sucre, Venezuela. Ciencia, 14(1), 12-27.
https://produccioncientificaluz.org/index.php/ciencia/article/view/9306
Mokhtari,
M., Ghaffar, M.A., Usup, F. y Cob, Z.C. (2016). Effects of fiddler crab burrows
on sediment properties in the mangrove mudflats of Sungai Sepang, Malaysia.
Biology, 5(7), 1-12. https://doi.org/10.3390/biology5010007
Moreno, C.E. (2001). Métodos para medir la biodiversidad. M y T SEA, Vol. 1. http://entomologia.rediris.es/sea/manytes/metodos.pdf
Negromonte, A.O., M.S.L. Câmara de Araújo, y P.A. Coelho. (2012). Decapod crustaceans from a marine tropical mangrove ecosystem on the southern
western Atlantic, Brazil. Nauplius, 20(2), 247-256. https://doi.org/10.1590/s0104-
64972012000200012
NOAA. Cold and warm episodes by season. Climate Prediction Center Database. https://origin.cpc.ncep.noaa.gov/products/analysis_monitoring/ensostuff/ONI_v 5.php
Olson, A.A. (1961). Panamic-Pacific pelecypoda. Mollusks of the tropical eastern Pacific. Particularly from the southern half of the panamic-pacific faunal province (Panama to Peru). Paleontological Research Institution. https://doi.org/10.5962/bhl.title.6853
Orellana Amador, J.J. (2010). Peces, crustáceos y moluscos del istmo
centroamericano. OSPESCA. https://climapesca.org/wp-
content/uploads/2018/09/Peces,%20Crustaceos%20y%20Moluscos%20del%20Is tmo%20Centroamericano.pdf
Pan, F., K Xiao, A. Guo y H. LI. (2022). Effects of fiddler
crab bioturbation on the geochemical
migration and bioavailability of heave metals in coastal wetlands. J. Haz. Mat., 437, 129380. https://doi.org/10.1016/j.jhazmat.2022.129380.
Penha-Lopes,
F., F. Bartolini, S. LImbu, S. Camicci, e. Kristensen y J. Paula. (2009). Are fiddler crabs potentially useful ecosystem
engineers in mangrove wastewater wetlands? Mar. Poll. Bull., 58(11), 1694-1703.
https://doi.org/10.1016/j.marporbul.2006.06.015
Pfaff, M.C. y R. Nel. (2019). Intertidal zonation. Reference module in earth system and environmental sciences. Second Edition, Elsevier. 97-107.
https://doi.org/10.1016/B978-0-12-409548-9.11184-4
Pomareda, E. y I. Zanella. (2006). Mollusks diversity
associated with mangroves in San Lucas Island. Rev. Cienc. Amb., 32(1), 11-13. https://doi.org/10.15359/rca.32-1.3
Premcharoen, S., S. Witirawat y P. Tharapoom. (2016). Molluscan fauna in Bang Taboon mangrove estuary, inner Gulf of Thailand: Implications for conservation and sustainable use of coastal resources. MATEC Web Conf., 60, 1-5.
https://doi.org/10.1051/matecconf/20166002003
Quinceno Cuartas, P.A. y J.A. Palacio Baena. (2008). Aporte
al conocimiento de los macroinvertebrados
asociados a las raíces del mangle (Rhizophora mangle) en la ciénaga de la Boquilla, municipio de San Onofre, Sucre. Gestión y Ambiente, 11(3), 67-68. https://revistas.unal.edu.co/index.php/gestion/article/view/14021
Quirós,
J.A. y J.E. Arias. (2013). Taxocenosis de moluscos y crustáceos en raíces de Rhizophora
mangle (Rhizophoraceae) en la Bahía de Cispatá, Córdoba, Colombia. Acta Biológica Colombiana, 18(2), 329-340.
https://www.redalyc.org/pdf/3190/319028011009.pdf
Rahardjanto, A., Tosiyana, V. R., Husamah, H., Miharja, F. J. (2020). Diversity of mollusks in the mangrove forest area of Cengkrong beach-trenggalek. AIP Conference Proceedings, 2231(246):1–7. https://doi.org/10.1063/5.0002618
Rodríguez, E.A., J.C. Chang y I. Goti. (2012).
Productividad primaria del manglar de Rhizophora mangle L. en el canal
estuarino de Isla de Cañas, provincia de Los Santos, República de Panamá. Tecnociencia, 14(2), 85-99.
https://revistas.up.ac.pa/index.php/tecnociencia/article/view/1005
Ross, E., J.M. Posada, A. Piedra, J.M. Díaz y G. Melo. (2014). Guía de identificación: Invertebrados marinos de importancia comercial en la costa Pacífica de Costa Rica. MarViva. https://marviva.net/wp-
Saint-Paul, U. (2007). Interrelations among mangroves, the local economy and social sustainability: a review from a case study
in north Brazil. In Hoanh, C.Y., J.W. Tuiong
y B. Hardy (eds.). Environmental and livelihoods in tropical coastal zone. 154-162.
http://www.iwmi.cgiar.org/Publications/CABI_Publications/CA_CABI_Series/Coa stal_Zones/Hoanh_1845931076-Chapter12.pdf
Sanders, H. L. (1968). Marine benthic diversity: a comparative study. Am. Nat., 102(925), 243-282. https://www.journals.uchicago.edu/doi/epdf/10.1086/282541
Sánchez-Andrés, R., S. Sánchez-Carrillo, L.C. Alatorre, S.
Cirujano, M. Álvarez-Cobelas. (2010).
Litterfall dynamics and nutrient decomposition of arid mangroves in the Gulf of California: Their role sustaining
ecosystem heterotrophy. Est. Coast. Shelf Sci., 89(3), 191-199. https://doi.org/10.1016/j.ecss.2010.07.005
Satizabal, C.A., J.R. Cantera Kintz y P.C. Sierra-Correa.
(2012). Changes in mangrove epifaunal
assemblages caused by forest logging during hunting of the neotropical Cormorant (Phalacrocorax brasilianus) on
the Colombian Pacific coast. Open J. Mar. Sci.
2, 150-156. https://doi.org/10.4236/ojms.2012.24018
Setyadi, G., D.L. Rahayum, R.Pribadi, R. Harti, D.P. Wijayanti, D.N. Sugianto y A. Darmawan. (2021). Crustacean and mollusk species diversity and abundance in the mangrove communities of Mimika district, Papua, Indonesia. Biodiversitas, 22(10), 4146-4157. https://doi.org/10.13057/biodiv/d221004
Thurman, C.L., S.C. Faria y J.C. McNamara. (2013). The distribution of fiddler crabs (Uca) along the coast of Brazil: implications for biogeography of the
western Atlantic Ocean. Mar. Biodiv. Rec., 6(e1), 1-21.
https://doi.org/10.1017/S1755267212000942
Waran, M., M.I., Aipassa, J. Manusawai y A.S. Sinery.
(2020). Diversity of mollusks (gastropod and bivalve) in mangrove ecosystem of Oransbari district, south Manokwari Regency, west
Papua province, Indonesia. J. Environm.
Treat. Tech., 8(3), 1220-1224. https://doi.org/10.47277/8(3)1224


