INTRODUCCIÓN
Las comunidades de insectos acuáticos bentónicos poseen adaptaciones morfológicas, fisiológicas y conductuales que les permiten resistir las fuerzas de la corriente, sin embargo, una fracción de la comunidad bentónica, entra en la columna de agua y participan de un fenómeno conocido como deriva. Este fenómeno descubierto por Needham, (1928) y definido por Waters (1972) como el transporte de organismos macroinvertebrados, mayores a 0.30 mm aguas abajo, organismos bentónicos sumamente abundantes y diversos, en los ecosistemas acuáticos, lo que les confiere gran importancia para el entendimiento de la dinámica funcional, estructural y energética de estos ambientes (Aguirre-Pabón et al., 2012; Sánchez-Hernández, 2011).
Existen tres tipos de deriva: la deriva catastrófica, la deriva conductual y la deriva constante o causal (Water, 1962; Allan,1995), indistintamente al tipo de deriva al que se refiera, la dinámica del transporte de la fracción de los organismos bentónicos en la deriva, medido en términos de su abundancia, se convierte en un indicador de alteraciones en los ecosistemas acuáticos y por ende, en un indicativo de la salud ecológica o la calidad biológica del agua (Corigliano, et al.,1998; Sánchez-Hernández, 2011).
La periodicidad en la deriva exhibida por los organismos acuáticos, tanto en el trópico como en la zona templada, presenta un aumento en la abundancia de individuos durante el período nocturno y una reducción en su abundancia en el período diurno (Ramírez & Pringle, 1998; Quiñónez et al, 1998; Aguirre-Pabón et al., 2012; Tamaris-Turizo et al., 2013; Rodríguez & Duarte, 2019; Rodríguez & Otero, 2019). Este tipo de deriva llamada conductual o intencional por (Allan & Castillo, 2007), ocurre en respuesta a condiciones físicas, químicas o biológicas desfavorables o como una respuesta adaptativa inducida por la presión de los depredadores visuales, como los peces (Allan, 1995; Ramírez & Pringle 2001; Shearer et al., 2002).
Este mismo patrón en la periodicidad de la deriva fue encontrado por Flecker (1992) en ríos que históricamente presentan peces depredadores y una aperiodicidad en ríos que carecen de peces depredadores. La deriva nocturna como respuesta adaptativa a la depredación por parte de
peces, es apoyada por el hecho de que en la noche derivan las tallas más grandes de insectos, las cuales en el día serían más fácilmente detectadas por los depredadores visuales (Allan, 1984) y por el hecho de que los insectos que presentan deriva aperiódica son de tallas relativamente pequeñas, (Allan, 1978; Bello-González et al., 2015).
Sobre deriva de insectos acuáticos existe abundante literatura para zonas templadas (Brittain & Eikeland,1988;Ramírez & Pringle, 1998; Flecker, 1990, 1992; Pringle & Ramírez,1998), pero para la región Neotropical los estudios referentes a la composición, estructura y periodicidad son escasos y poco conocidos (Quiñones et al., 1998;Ramírez & Pringle, 1998, 2001; Rodríguez et al., 2006; Tamaris-Turizo, et al, 2013). Cabe destacar que en Panamá son muy limitados los registros de estudios sobre deriva, específicamente de insectos acuáticos (Rodríguez, 2018; Rodríguez & Duarte, 2019; Rodríguez & Otero, 2020), razón por la cual los objetivos principales de este estudio fueron: determinar la calidad del agua, la periodicidad de la deriva, la estructura numérica de la fracción de insectos acuáticos en la deriva, su relación con la comunidad bentónica de los insectos acuáticos en un tramo del río Zaratí, provincia de Coclé, Panamá.
MATERIALES Y MÉTODOS
El estudio se llevó a cabo en la parte baja del río Zaratí, específicamente en el balneario Las Mendozas, en el distrito de Penonomé, provincia de Coclé. El área ha sido clasificada según el índice bioclimático De Martonne, (1926), como un bioma per-húmedo. Se seleccionó un tramo, con una longitud de 40 m. y coordenadas 17 N 570802.3 y 942581.4
(UTM) 62 m s.n.m. (Fig. 1).
 Fig. 1.
Parte baja del
río Zaratí, balneario Las Mendozas,
Penonomé, República de Panamá.
Fig. 1.
Parte baja del
río Zaratí, balneario Las Mendozas,
Penonomé, República de Panamá.
El tramo fue seleccionado tomando en cuenta la mayor diversidad de hábitats posibles, para utilizar la técnica de muestreo de multi- hábitats, propuesta por Sermeño et al., (2010), para la recolecta de insectos acuáticos bentónicos. El muestreo se llevó a cabo entre los meses comprendidos entre junio y septiembre del 2019, dando un total de 12 campañas de muestreo. El esfuerzo de muestreo en el tramo estudiado fue de una hora para dos personas.
Método de recolección de los insectos acuáticos en la deriva.
Para la recolecta de insectos acuáticos en la deriva, se colocaron dos
trampas de deriva al inicio del tramo, cada una con un área de 0,176 m2
y una apertura de ojo de malla de 500 µm, donde se mantuvieron siempre
sumergidas y contra corriente. Las trampas de deriva se desocuparon cada 12
horas, durante un ciclo de 24 horas por día de colecta; (06:00 a 18: 00 h. y de
18:00 a 06:00 h.). Su contenido era depositado en bolsas “ziploc”, rotuladas y fijado con alcohol al 70% con cuatro gotas de glicerina.
Método de recolección de los insectos
acuáticos en la comunidad bentónica
La recolecta de los insectos acuáticos bentónicos se realizó, aguas abajo de las trampas de deriva y se utilizó una red tipo D-Net de 500 µm de
ojo de malla, con la que se hicieron barridos en los bordes, fondos, corrientes y remansos, procurando recolectar a los insectos acuáticos
desde aguas abajo hacia aguas arriba evitando no intervenir con las trampas de deriva.
El contenido de la red, se vaciaba, en una bandeja de color blanco y luego “in situ” con ayuda de pinzas entomológicas se extraían los insectos acuáticos. Para registrar la mayor cantidad de taxa, se utilizaron también pinzas entomológicas para recolectar los organismos directamente del sustrato. Todos los insectos acuáticos recolectados fueron almacenados en envases plásticos de 150 ml., rotulados y fijados en alcohol al 70%, más dos gotas de glicerina para mantener blandas y flexibles las estructuras de los organismos. Todo el material recolectado fue transportado al laboratorio del Centro Regional Universitario de Coclé, Universidad de Panamá.
Tratamiento de muestras en laboratorio
Los insectos acuáticos fueron cuantificados e identificados hasta el nivel de género empleando
las claves taxonómicas de Novelo-Gutiérrez, (1997a, 1997b); Roldán-Pérez
(1988); Flowers & De La Rosa (2010); Ramírez (2010); Gutiérrez-Fonseca (2010); Springer (2006, 2010).
Análisis de la calidad biológica del agua: Índice EPT
El análisis EPT se realizó mediante la utilización de estos tres órdenes de
insectos acuáticos (Ephemeroptera, Plecoptera y Trichoptera) que son
indicadores de buena calidad de agua, debido a su alta sensibilidad a la contaminación. Para ello, se utilizó el método llamado EPT
(%), el que se obtuvo dividiendo el número de individuos de estos órdenes
presentes, entre el número total de individuos recolectados; el valor obtenido
se multiplicó por cien y el resultado se comparó en una tabla de calificaciones
de calidad de agua (Carrera y Fierro, 2001).
Análisis estadístico
Se aplicaron las pruebas de Mann-Whitney y Friedman, para comparar abundancia, y para determinar diferencias, se utilizó la prueba de comparaciones múltiples por pares, mediante el procedimiento de Steel- Dwass-Critchlow-Fligner. La diversidad se midió con el índice de diversidad de Shannon-Weaver (H’) y se utilizó la t de Hutcheson, para determinar diferencias. El coeficiente de similitud de Jaccard, fue utilizado para medir la similitud entre las comunidades. Los datos fueron analizados en el software Past 3.16 (Hammer, Harper y Ryan,
2001) para Windows y en todas las pruebas estadísticas se utilizó un α
≤ 0.05.
RESULTADOS
Estructura numérica de la comunidad
bentónica de insectos acuáticos en el tramo estudiado del río Zaratí.
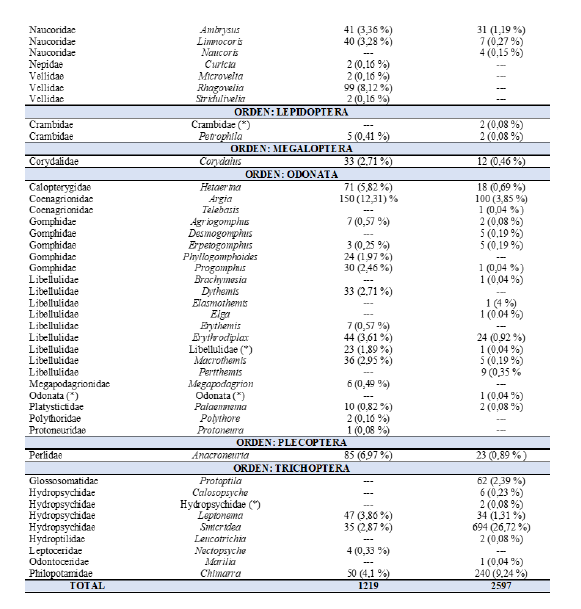
En el Cuadro 1, se presentan los resultados de los órdenes, familias y géneros de insectos acuáticos, en la comunidad bentónica, reportados en Cárdenas, (2017). Se encontraron diferencias significativas en la abundancia de los insectos acuáticos entre el presente estudio y el de Rodríguez & Cárdenas, (2017), (Friedman, Q= 9,87, gl= 2, p= 0.007),
(Cuadro 1).
Cuadro 1.
Familias y géneros reportados por Rodríguez y Cárdenas
(2017) en comparación con el presente estudio.
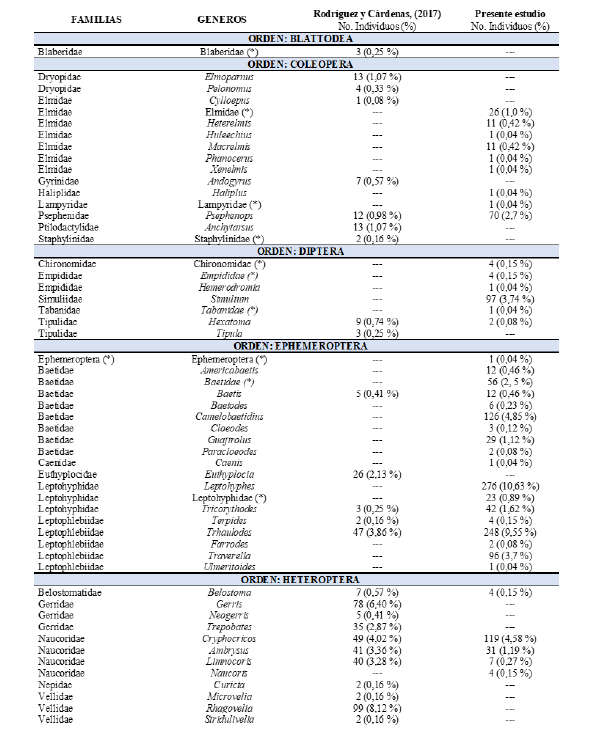
En el tramo estudiado del río Zaratí se recolectaron un total de 2 597 insectos acuáticos en la comunidad bentónica, distribuidos en 9 órdenes, 28 familias (dos familias sin determinar) y 56 géneros (12 géneros sin determinar). En términos de mayor a menor abundancia los órdenes se distribuyeron de la siguiente manera: Trichoptera con 1 041 individuos, lo que representa el (40,08 %) de la comunidad bentónica de insectos acuáticos, seguido de Ephemeroptera con 940 (36,2 %), Odonata con 177 (6,82 %), Heteroptera con 168 (6,47 % ), Coleoptera con 123 (4,74
% ), Diptera con 109 (4,2 % ), Plecoptera con 23 (0,89 % ), Megaloptera
con 12 (0,46 % ) y Lepidoptera 4 (0,15 % ), (Fig. 2 y 6, Cuadro1).
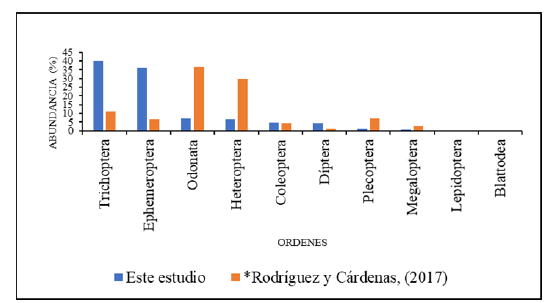 Fig. 2.
Porcentaje de abundancia de los órdenes de insectos acuáticos en la comunidad bentónica, en el tramo estudiado del río Zaratí, comparado
con el estudio de Rodríguez y Cárdenas, (2017).
Fig. 2.
Porcentaje de abundancia de los órdenes de insectos acuáticos en la comunidad bentónica, en el tramo estudiado del río Zaratí, comparado
con el estudio de Rodríguez y Cárdenas, (2017).
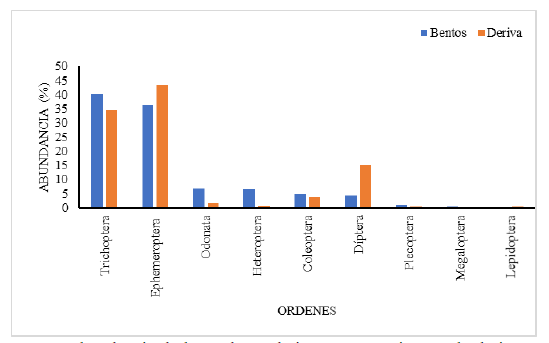 Fig. 3.
Abundancia de los órdenes de insectos
acuáticos en la deriva y en la comunidad bentónica, en el tramo estudiado del
río Zaratí.
Fig. 3.
Abundancia de los órdenes de insectos
acuáticos en la deriva y en la comunidad bentónica, en el tramo estudiado del
río Zaratí.
Se encontró diferencias significativas en la abundancia del grupo EPT (Ephemeroptera, Plecoptera y Trichoptera) entre Rodríguez y Cárdenas (2017) y el presente estudio (Mann-Whitney, U Test, p= 0,000), (Fig, 3). El índice EPT calculado en el presente estudio fue de 68,71 ± 14 (n
= 12), lo que indica una calidad biológica del agua, como muy buena.
Se encontraron en el tramo estudiado del río Zaratí, del orden Ephemeroptera, los géneros: Baetodes, Caenis, Camelobaetidius, Cloeodes, Farrodes, Guajirolus, Leptohyphes, Leptohyphidae (sin determinar), Paracloeodes, Baetidae (sin determinar), Traverella, Americabaetis, Baetidae (sin determinar) y Ulmeritoides, géneros no encontrados por Rodríguez y Cárdenas (2017). Del orden Trichoptera encontramos los géneros Calosopsyche, Hydropsychidae (sin determinar), Leucotrichia, Marilia y Protoptila los cuales no fueron reportados por Rodríguez y Cárdenas (2017), (Cuadro 1).
Los órdenes Heteroptera,Odonata, Trichptera y Ephemeroptera mostraron diferencias significativas en su abundancia, entre este estudio y el realizado por Rodríguez y Cárdenas (2017) (Mann-Whitney, U-test, p < 0,05. (Fig. 2). Dentro del orden Heteroptera, los géneros: Curicta, Gerris, Neogerris, Rhagovelia, Stridulivelia y Trepobates, y los géneros: Dythemis y Erythemis del orden Odonata no fueron colectados en el presente estudio. De los 89 géneros totales encontrados en ambos estudios el 55 % de los géneros corresponden a los reportados por Rodríguez y Cárdenas, (2017) y 76,4 % a los reportados en este estudio.
Las familias más abundantes de insectos acuáticos en la comunidad bentónica del tramo estudiado en el río Zaratí fueron: Hydropsychidae, Leptophlebiidae, Leptohyphidae, Baetidae, Philopotamidae, Naucoridae, Coenagrionidae, Simuliidae, Psephenidae, Glossosomatidae y Elmidae las que representan el 94,57 % de los individuos (Fig. 3), mientras que, las menos abundantes fueron: Libellulidae, Perlidae, Calopterygidae, Gomphidae, Corydalidae, Belostomatidae, Chironomidae, Empididae, Crambidae, Tipulidae, Platystictidae, Hydroptilidae, Haliplidae, Lampyridae, Empididae, Tabanidae, Caenidae, Ephemeroptera (sin determinar), Odonata (sin determinar) y Odontoceridae quienes representaron el 5,43 % del total de los individuos (Fig. 3. como otras).
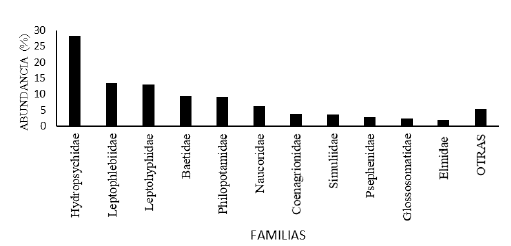 Fig. 4.
Porcentaje de abundancia de las familias de insectos acuáticos en la comunidad bentónica, en el tramo estudiado del río Zaratí, provincia de Coclé, Panamá.
Fig. 4.
Porcentaje de abundancia de las familias de insectos acuáticos en la comunidad bentónica, en el tramo estudiado del río Zaratí, provincia de Coclé, Panamá.
En el tramo estudiado del río Zarati,
los géneros de insectos acuáticos más abundante en la comunidad bentónica
fueron: Smicridea,
Leptohyphes,
Thraulodes,
Chimarra, Camelobaetidius, Cryphocricos, Argia, Simulium, Traverella, Psephenus, Protoptila, Tricorythodes,
Baetidae (género sin determinar), Leptonema, Ambrysus, Guajirolus, Elmidae
(género sin determinar) y Erythodiplax, que representaron el 90,64 % del total de
individuos recolectados (Fig. 4).
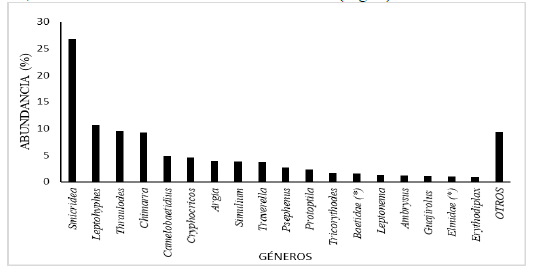 Fig. 5.
Abundancia porcentual de los géneros de
insectos acuáticos en la comunidad bentónica, en el tramo estudiado del río Zaratí, provincia de Coclé, Panamá.
Fig. 5.
Abundancia porcentual de los géneros de
insectos acuáticos en la comunidad bentónica, en el tramo estudiado del río Zaratí, provincia de Coclé, Panamá.
Mientras que, los géneros menos abundantes fueron: Leptohyphidae (género sin determinar), Anacroneuria, Hetaerina, Prebaetodes, Baetis, Americabaetis, Corydalus, Heterelmis, Macrelmis, Perithemis, Limnocoris, Baetodes, Calosopsyche, Erpetogomphus, Desmogomphus, Macrothemis, Chironomidae (género sin determinar), Empididae (género sin determinar), Terpides, Belostoma, Naucoris, Cloeodes, Lethocerus, Hexatoma, Paracloeodes, Farrodes, Petrophila, Crambidae (género sin determinar), Agriogomphus, Palaemnema, Hydropsychidae (género sin determinar), Leucotrichia, Phanocerus, Xenelmis, Huleechius, Haliplus, Lampyridae (género sin determinar), Hemerodromia, Tabanidae (género sin determinar), Caenis, Ulmeritoides, Ephemeroptera (género sin determinar), Telebasis, Progomphus, Brachymesia, Libellulidae (género sin determinar), Elasmothemis, Elga, Odonata (género sin determinar) y Marilia quienes representaron el 9,36 % de los individuos (Fig. 4, como otros).
Periodicidad en la deriva de insectos acuáticos en el tramo estudiado del río Zaratí.
Durante las 12 giras de muestreos, en el tramo estudiado del río Zaratí se recolectaron un total de 1 387 insectos acuáticos en la deriva, 206 individuos en el período diurno de 06:00 a 18: 00 h y 1 181 individuos en el periodo nocturno de 18:00 a 06:00 h. Se encontraron diferencias significativas en la abundancia de los insectos acuáticos presentes en la deriva entre el período diurno y nocturno, (Friedman, Q= 3,8, gl= 1, p= 0,0001). La prueba de comparaciones múltiples por pares, mediante el procedimiento de Steel-Dwass-Critchlow-Fligner / Prueba bilateral; diferenció dos grupos de medianas homogéneas (06:00 a 18: 00 h.) y (18:00 a 06:00 h.). La mayor abundancia de insectos en la deriva en el tramo estudiado del río Zaratí, se registró en el período nocturno de 18:00 a 06:00 h., (Fig. 5).
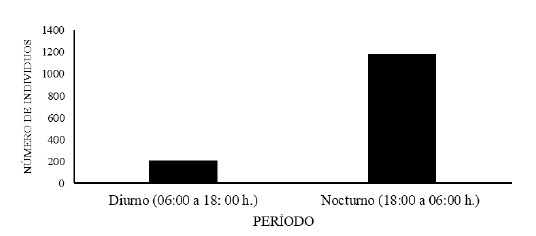 Fig. 6.
Abundancia de insectos acuáticos en la deriva en los distintos periodos de muestreo, en el
tramo estudiado del río Zaratí, Coclé, Panamá.
Fig. 6.
Abundancia de insectos acuáticos en la deriva en los distintos periodos de muestreo, en el
tramo estudiado del río Zaratí, Coclé, Panamá.
El cálculo de la diversidad fue de H’= 2,66 (J’=0,81 Taxa=
27) para el período de diurno de 6:00 -18:00 h. y de H’= 2,61 (J’=0,66 Taxa= 51) para el período nocturno de 18:00-6:00 h. No se encontraron diferencias significativas en la
diversidad para los distintos periodos de muestreo (p=0,928; t de Hutcheson).
Estructura numérica de los insectos
acuáticos en la deriva, en el tramo estudiado del río Zaratí.
De forma general, la entomofauna acuática en la deriva estuvo agrupada en 9 órdenes; 30 familias, 43 géneros determinados y 10 géneros sin determinar. En términos de abundancia, los órdenes se distribuyeron de la siguiente manera: Ephemeroptera con 602 individuos, lo que representa el (43,4 %), Trichoptera con 478 (34,46 %), Diptera con 209
(15,07 %), Coleoptera con 52 (3,75 %), Odonata con 22 (1,59
%), Heteroptera con 10 (0,72 %), Lepidoptera con 6 (0,43 %),
Plecoptera con 5 (0,36 %) y Megaloptera con 3 (0,22 %), (Fig. 6).
Las familias más abundantes en la fracción de insectos en la deriva, en el tramo estudiado del río Zaratí fueron: Hydropsychidae, Baetidae, Leptohyphidae, Chironomidae , Hydroptilidae, Leptophlebiidae, Elmidae, Tabanidae, Haliplidae, Naucoridae, Coenagrionidae y Libellulidae, las cuales representaron el 96,76 % de los individuos (Fig. 7), mientras que, las menos abundantes fueron: Ephemeroptera (familia sin determinar), Crambidae, Perlidae, Simuliidae, Ceratopogonidae,
Corydalidae, Hydrophilidae, Scirtidae, Oligoneuriidae, Calopterygidae, Gomphidae, Glossosomatidae, Lampyridae, Psephenidae, Empididae, Caenidae, Leptoceridae y Odontoceridae familias que representaron el 3,24 % (Fig. 7, como otras).
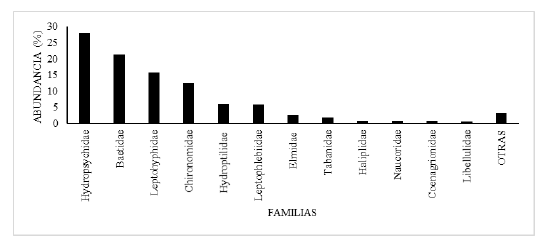 Fig. 7.
Porcentaje de abundancia de las familias de insectos acuáticos en la fracción derivante, de la comunidad bentónica, en el tramo estudiado del río Zaratí.
Fig. 7.
Porcentaje de abundancia de las familias de insectos acuáticos en la fracción derivante, de la comunidad bentónica, en el tramo estudiado del río Zaratí.
En la fracción de insectos acuáticos en la deriva, los géneros más abundantes fueron: Smicridea, Leptohyphes, Chironomidae (sin determinar), Camelobaetidius, Leucotrichia, Baetidae (*), Guajirolus, Thraulodes, Terpides, Tabanidae (*), Americabaetis, Baetidae (*), Tricorythodes, Elmidae (*), Leptohyphidae (*), Paracloeodes, Leptonema, y Haliplus géneros que representaron el 91,56 % de la deriva (Fig. 8). Los géneros menos abundantes fueron: Macrelmis, Baetis, Cloeodes, Traverella, Ephemeroptera (sin determinar), Cryphocricos, Petrophila, Heterelmis, Farrodes, Anacroneuria, Phanocerus, Simulium, Stilobezzia, Baetodes, Naucoris, Corydalus, Erythodiplax, Macrothemis, Hydrophilidae (sin determinar), Elodes, Lachlania, Hetaerina, Epigomphus, Perithemis, Protoptila, Hexanchorus, Lampyridae (sin determinar), Psephenus, Empididae (sin determinar), Callibaetis, Caenis, Ambrysus, Neptopsyche y Marilia, géneros que representan el 8, 44 % de la deriva (Fig. 8, como otros).
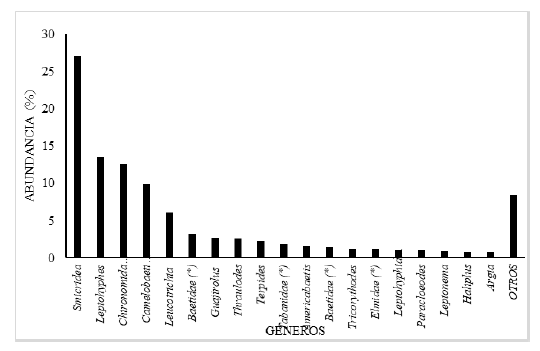 Fig. 8.
Porcentaje de abundancia de los géneros de insectos acuáticos en la fracción derivante, de la comunidad bentónica, en el tramo estudiado del río Zaratí.
Fig. 8.
Porcentaje de abundancia de los géneros de insectos acuáticos en la fracción derivante, de la comunidad bentónica, en el tramo estudiado del río Zaratí.
Relación entre la comunidad bentónica
de insectos acuáticos y la fracción de insectos acuáticos en la deriva.
El cálculo de la diversidad fue de H’= 2,66 (J’= 0,67, Taxa= 53) para la fracción de insectos acuáticos en la deriva y de H’= 2,8 (J’= 0,66, Taxa= 68) para la comunidad bentónica de insectos acuáticos. Se encontraron diferencias significativas en la diversidad entre la fracción en la deriva de insectos acuáticos y la comunidad bentónica en el tramo estudiado. La diversidad en la comunidad bentónica fue mayor que en la fracción de insectos en la deriva (p= 0,002; t de Hutcheson).
De los 53 géneros de insectos acuáticos capturados en las redes de deriva, 49 géneros (92,4 %) están presentes en la comunidad bentónica, del tramo estudiado, mientras que 26 (38,2 %) de los 68 géneros presentes en la comunidad bentónica no se capturaron en las redes de deriva. En la fracción de insectos acuáticos en la deriva se encontraron 8 (14,8 %) géneros, que no se capturaron en la comunidad bentónica del tramo estudiado del río Zaratí. El índice de similitud de Jaccard mostró un valor de similitud entre la comunidad bentónica y la fracción de insectos acuáticos en la deriva de IJ= 52 %.
DISCUSIÓN
Comparación entre la estructura
numérica de la comunidad bentónica de insectos acuáticos reportada por
Rodríguez y Cárdenas, (2017) y la estructura numérica en el encontrada en el
presente estudio, en el tramo estudiado del río Zaratí.
Rodríguez y Cárdenas (2017), registran en la comunidad bentónica del río Zaratí, 10 órdenes de insectos acuáticos, que coinciden con el presente estudio a excepción del orden Blattodea, que no fue colectado en este estudio. Según Rodríguez y Cárdenas (2017), los órdenes más abundantes, en tres tramos estudiados del río Zaratí fueron: Odonata, Heteroptera, Trichoptera, y Ephemeroptera los que representaron el 84,46 % de los insectos acuáticos recolectados, mientras que los más abundantes en este estudio fueron: Trichoptera, Ephemeroptera, Odonata y Heteroptera los que representaron el 89,56 %, mismos órdenes que los encontrados por Rodríguez y Cárdenas (2017) como los más abundantes a excepción del orden Plecoptera.
Al encontrar diferencias significativas en la abundancia de los órdenes Trichoptera, Ephemeroptera, Odonata, Heteroptera y Trichoptera, entre Rodríguez y Cárdenas (2017) y el presente estudio, se debe principalmente a la aparición en este estudio de los géneros del orden Ephemeroptera: Baetodes, Caenis, Camelobaetidius, Cloeodes, Farrodes, Guajirolus, Leptohyphes, Leptohyphidae (sin determinar), Baetidae (sin determinar), Traverella, Americabaetis, Baetidae (sin determinar) y Ulmeritoides, géneros no encontrados por Rodríguez y Cárdenas (2017). También encontramos géneros del orden Trichoptera como: Calosopsyche, Hydropsychidae (sin determinar), Leucotrichia, Marilia y Protoptila los cuales no fueron encontrados por Rodríguez y Cárdenas (2017). El índice EPT (%) calculado en el presente estudio fue de 68,71 ± 14 (n = 12), lo que indica una calidad biológica del agua, como muy buena, mientras que el calculado por Rodríguez y Cárdenas (2017) fue de 22,5 ± 14,2 (n=30), lo que indica una mala calidad de agua. La diferencia en la calidad biológica del agua, entre el estudio de Rodríguez y Cárdenas, (2017) y el presente estudio podría deberse al incremento de los géneros de Ephemeroptera, Plecoptera y Trichoptera (EPT) en el presente estudio y a la disminución en la abundancia de los órdenes Heteroptera y Odonata, órdenes abundantes en ríos con algún
grado de perturbación en la provincia de Veraguas (Rodríguez & Sánchez,
2001,Rodríguez & Mendoza, 2003,Lombardo & Rodríguez, 2007). Otra posibilidad que explicaría el cambio en la calidad biológica del agua podría deberse a que el río Zaratí
al encontrarse en un bioma per-húmedo (De Martonne, 1927), con registros
históricos de precipitación de 3 738,8 mm al año (n=63 años) (ETESA, Open Data,
estación Chiriguí Arriba, código 105-002, 2016), las
fuertes lluvias podrían arrastrar la contaminación y a la vez, alterar las condiciones físico-químicas
del agua, y alterar la estructura de las comunidades bentónicas de insectos
acuáticos e influir en la calidad biológica del agua, ya que perturbaciones
naturales que incrementan la precipitación pueden afectar la química del agua (Schaefer et al., 2000; Blanco et al.,
2003).
Periodicidad en la deriva de insectos acuáticos en el tramo estudiado del río Zaratí.
Las mayores abundancias de insectos acuáticos en la deriva en el tramo estudiado del río Zaratí, se obtuvieron en el período nocturno (18:00- 06:00 h.). Se ha documentado, en Panamá patrones en la periodicidad de la deriva de insectos acuáticos, mostrando una deriva baja y constante durante el día, seguida por incrementos en el número de organismos durante la noche (Guerra, 2017; Tuñón, 2019; Rodríguez & Otero, 2020; Castillo & Rodríguez, 2020). En otras regiones también se ha documentado el mismo patrón en la periodicidad en la deriva (Waters, 1962; Ramírez & Pringle, 1998;Aguirre-Pabón, et al., 2012; Tamaris- Turizo, et al., 2013). Este tipo de deriva es llamada “conductual” por Allan y Castillo, (2007), ocurre generalmente en respuesta a condiciones desfavorables o responde a comportamientos evasivos ante el riesgo de depredadores (Shearer, et al., 2002). Estudios en la periodicidad de la deriva, en ríos de los Andes venezolanos donde históricamente no han existido depredadores, presentaron un patrón de deriva aperiódico (Flecker, 1992) y los insectos que presentaron deriva aperiódica son relativamente más pequeños, (Allan, 1978; Bello-González et al., 2015) parece apoyar la “hipótesis de deriva de insectos acuáticos como un mecanismo para evitar la depredación”.
Estructura numérica de los insectos
acuáticos en la deriva, en el tramo estudiado del río Zaratí.
En el tramo estudiado del río Zaratí, la caracterización de la
fracción de
insectos acuáticos en la deriva, en términos de abundancia, se encontró dominada por los órdenes: Ephemeroptera, Trichoptera, Díptera y Coleoptera, los que representaron el 96,7 % de la colecta. En la provincia de Veraguas los estudios realizados por Guerra (2017), Aparicio & Concepción (2017), Rodríguez & Duarte, (2019) y Rodríguez & Otero, (2010) reportan como los órdenes de insectos más abundantes en la deriva a: Ephemeroptera, Trichoptera, Díptera y Coleoptera, con porcentajes de abundancia de: 90,9 %; 94,4 %; 93,8 %; y 92,1 % respectivamente, coincidiendo con el presente estudio, lo que parece indicar que los órdenes: Ephemeroptera Trichoptera, Coleoptera y Diptera son los órdenes más propensos a desplazarse en la deriva. Tuñón (2019), en Veraguas y Castillo & Rodríguez (2020), en la provincia de Herrera, reportan a los órdenes: Ephemeroptera Trichoptera, Coleoptera y Plecoptera con una abundancia del 82,6 % y el 91 % respectivamente, de insectos acuáticos en la deriva, órdenes que coinciden con el presente estudio, como los órdenes más propensos a entrar en la deriva, a excepción del orden Plecoptera.
Estudios realizados en la fracción derivante de insectos acuáticos, concernientes a otras regiones neotropicales, como los de Aguirre- Pabón et al., (2012) documentaron, en términos generales, a los órdenes Diptera y Ephemeroptera como los que tienen mayor tendencia a derivar, y representaron el 93,4 % de la deriva. Callisto & Goulart, (2005) encontraron que los órdenes Diptera, Trichoptera y Ephemeroptera son los más propensos a entrar en la deriva y representaron el 87.9 % de la deriva total. Tamaris-Turizo et al., (2013), reportaron que los órdenes Diptera, Ephemeroptera y Trichoptera, son los que presentaron las mayores densidades en la deriva; mientras que, Rodríguez-Barrios et al., (2007), reportaron que los órdenes Diptera, Trichoptera y Ephemeroptera con las mayores densidades en la deriva. Ordenes que coinciden con el estudio como los más propensos a entrar en la deriva, a excepción de Coleoptera, que en el presente estudio se encuentra como un orden propenso a entrar en la deriva.
Relación entre la comunidad bentónica de insectos acuáticos y la fracción de insectos acuáticos en la
deriva.
Al caracterizar la comunidad bentónica, en términos de abundancia, en el tramo estudiado del río
Zaratí, los órdenes más abundantes fueron: Trichoptera, Ephemeroptera, Odonata y Heteroptera los que
representaron el 89,77 % de la abundancia, mientras que, la caracterización de la comunidad de insectos acuáticos en la deriva, en términos de abundancia, se encontró dominada por los órdenes: Ephemeroptera, Trichoptera, Díptera y Coleoptera. Aunque parece no existir un patrón en la composición taxonómica en cuanto a órdenes de insectos acuáticos en la deriva, todo parece indicar que los órdenes de insectos acuáticos más abundantes en la comunidad bentónica, corresponden a los mismos órdenes abundantes en la deriva. Matthaei, et al., (1998) y Ramírez & Pringle, (1998) reportaron que los órdenes con mayor abundancia en la deriva, son los mismos órdenes abundantes de la comunidad bentónica y pertenecen a los órdenes Diptera, Trichoptera, Ephemeroptera y Plecoptera. Ramírez & Pringle (1998) documentan que la deriva y la comunidad bentónica están compuestos por grupos similares, pero con diferentes proporciones de abundancia. Los órdenes que dominan en abundancia, la fracción derivante, coinciden con los órdenes más abundantes en las comunidades bentónicas (Ríos, 2008).
El porcentaje de géneros de insectos acuáticos en la deriva, que están presente en la comunidad bentónica, en este estudio fue de 92,4 %, mientras que, Aparicio & Concepción (2017) y Rodríguez & Duarte (2019), reportan porcentajes de 70 %, y 90,4 %, respectivamente. El elevado porcentaje de géneros de insectos acuáticos de las comunidades bentónicas, capturados en la fracción derivante, indica que la mayor parte de los géneros de las comunidades bentónicas se desplazan en la deriva. Bello-González et al., (2015) encontraron en el río Bayate (Cuba), que el 74,3 % de los géneros encontrados en la comunidad bentónica fueron hallados también participando en la deriva. Sánchez- Hernández, (2011) comunica para el río Tormes (España) que los géneros más abundantes en la comunidad bentónica están representados en la deriva en mayor o menor medida. Es cierto, que todos los géneros de insectos de la comunidad bentónica tienen la misma posibilidad de entrar en la deriva, pero no todos tienen la misma predisposición a derivar, y por lo tanto la composición de la fracción de insectos en la deriva no sería exactamente igual al de la comunidad bentónica. Los géneros de los órdenes de Ephemeroptera, Trichoptera, Diptera y Coleoptera tienen mayor posibilidad de ser transportados en la deriva, mientras que, los géneros de los órdenes de Lepidóptera, Plecoptera, Heteroptera, Odonata, Megaloptera y Blattodea, son los menos
propensos a derivar (Elliott, 1967,Corigliano et al.,1987).
El índice de similitud de Jaccard mostró un valor de similitud entre los géneros encontrados en la fracción de insectos acuáticos en la deriva y los géneros encontrados en la comunidad bentónica, en el tramo estudiado del río Zaratí de IJ= 52 %. Mientras que, Rodríguez & Duarte, (2019) reportan un índice de similitud de IJ= 75 %, para el tramo estudiado del río Santa María. Las semejanzas entre las comunidades bentónicas y la fracción derivante, indica que la deriva es una expresión de la estructura de la comunidad bentónica, en concordancia con la hipótesis de denso-dependencia. Similar a lo comunicado por Gualdoni & Corigliano, (1999), y Zanotto-Arpellino et al., (2015) al encontrar una elevada similitud entre la estructura de la comunidad bentónica y de la fracción de insectos en la deriva, de 75 % y 86 % respectivamente. Por lo que las altas tasas de transporte en la deriva de los grupos dominantes corresponden a sus altas abundancias en la comunidad bentónica (Ríos, 2008).
La importancia de la densidad bentónica, como uno de los principales factores que inducen a los insectos a derivar, ha sido estudiada por diferentes autores, las conclusiones no siempre fueron concordantes. Mientras que, algunos autores encuentran una relación entre la fracción derivante y la densidad bentónica (Allan, 1978, Gualdoni & Corigliano, 1999, Zanotto-Arpellino et al., 2015 ), otros consideran que las dos fracciones presentan diferentes patrones de abundancia (Elliott, 1967, Corkum, 1978, Koetsier & Bryan, 1996,Principe & Corigliano, 2006), lo que parece indicar que son múltiples los factores que pueden influir sobre este fenómeno, y la densidad bentónica no es el único disparador (Gualdoni & Corigliano, 1999).
CONCLUSIONES
Las características estructurales de la fracción de insectos acuáticos en la deriva y la caracterización de las comunidades bentónicas, unido a las semejanzas entre las comunidades bentónicas y la fracción de insectos en la deriva, parece indicar al menos que, la deriva es una expresión de la estructura de la comunidad bentónica.
La tendencia a derivar, además de la densidad bentónica, puede estar asociada también, con algunos aspectos de la forma de vida de los organismos que integran la fracción derivante, como los son: la forma corporal hidrodinámica que determina la habilidad para nadar, las adaptaciones morfológicas y etológicas que favorecen la permanencia en el sustrato, el microhábitat preferencial y la velocidad de la corriente.
Los valores contrastantes en la abundancia de insectos acuáticos en la deriva, altos en el periodo nocturno y bajos en el período diurno, sugieren que la deriva en el tramo estudiado es periódica y fundamentalmente conductual e intencional. Los individuos entran a la columna de agua generalmente como parte de su comportamiento y no por accidente, lo que parece reducir el riesgo de depredadores visuales diurnos durante la movilización en la deriva.
REFERENCIAS
Aguirre-Pabón, J. Rodríguez-Barrios, J. & Ospina-Torres, R. (2012). Deriva de macroinvertebrados acuáticos en dos sitios con diferente grado de perturbación, río Gaira, Santa Marta-Colombia. Intropica. Vol. 7, (1), págs. 9-19.
Allan, J. D. 1978. Trout predation and the size composition of stream drift. Limnology and Oceanography. 23(6), 1231-1237.
Allan, J. D. 1984. The size composition of invertebrate drift in a Rocky Mountain stream. Oikos. 43, 68-76.
Allan, J.D. (1995). Stream Ecology: Estructure and function of running waters. Edit. Chatman y Hall. 388 pp.
Allan, J. D. & Castillo, M. M. (2007). Detrital energy sources. En J. Allan Y M. M. Castillo (Eds.). Stream ecology structure and function of running waters. (pp.135–161). Netherlands: Springer Dordrecht.
Bello-González, O. C., Curbelo, G. E., Fontenla, Y., Botello, F. D., Castillo, I. de la C., Santalla, M., & Benítez, R. (2015). Deriva de macroinvertebrados acuáticos en un afluente del río Bayate, Sierra del Rosario, Cuba. Revista Cubana de Zoología. Poeyana. (501), 1-7.
Blanco, J.; Vásquez, G.; Ramírez, J, & Bavarrete, A. M. (2003). Variación de algunos parámetros fisicoquímicos en el río Pescador, Valle del Cauca, durante el ciclo El Niño 1997/1998 y La Niña 1998/1999. Revista Actualidades Biológicas. 25(78):59-69.
Brittain, J. E., & Eikelands, T. J. (1988). Invertebrate drift—a review. Hydrobiologia 166: 77–93.
Callisto, M. & Goulart, M. (2005). Invertebrate drift along a longitudinal gradient in a Neotropical stream in Serra do Cipo´ National Park, Brazil. Hydrobiologia (539), 47–56.
Carrera, C. y Fierro, K. (2001). Manual de monitoreo los macroinvertebrados acuáticos como indicadores de la calidad del agua. Fundación EcoCiencia. Quito. Ecuador.
Castillo, M. & Rodríguez, M. (2020). Estructura numérica de la fracción derivante de insectos acuáticos y su relación con el béntos en la reserva forestal el montuoso, distrito de Las Minas, provincia de Herrera. Centro Regional Universitario de Veraguas, Facultad de Ciencias Naturales Exactas y Tecnología. Tesis de Grado. Biología. Panamá.
Corigliano, M. C.; Gualdoni, A. M.; Oberto & Raffaini, G. B. (1998). Atributos estructurales de la deriva de invertebrados en el río Chocancharava, Córdoba, Argentina. Ecología Austral, (8), 5-12.
Corigliano, M. C.; Gualdoni, C. M. & Oberto, A. M. (1987). Deriva de macroinvertebrados en un tramo anastomosado de un río de llanura. Revista Universidad Nacional de Rio Cuarto 7(1):89-98.
Corkum, L. D., (1978). The influence of density and behavioural type on the active entry of two mayfly species (Ephemeroptera) into the water column. Canadian Journal of Zoology 56(5):1201-1206.
De Martonne, E. (1927). Regions of Interior-Basin Drainage. Geographical Review, 17(3), pp. 397-414. doi:10.2307/208323
Elliott, J. M. (1967). The life histories and drifting of the Plecoptera and Ephemeroptera in a Dartmoor stream. Journal of Animal Ecology. Vol. (36), 343-362.
ETESA (2016). Open Data recuperado el 28 de julio de 2021 de https://www.hidromet.com.pa/es/clima-historicos
Flecker, A.S. (1992), Fish Predation and The Evolution of Invertebrate Drift Periodicity: Evidence from Neotropical Streams. Ecology, 73: 438-448. https://doi.org/10.2307/1940751
Flowers, R.W. & De la Rosa, C. (2010). Capítulo 4: Ephemeroptera. Revista Biología Tropical. Vol. 58 (Suppl.4): 63-93.
Gualdoni, C. M. y Corigliano, M. C. (1999). Deriva de insectos y su relación estructural con el bentos. Idesia (17), 57-71.
Guerra, B. (2017). Densidad y estructura numérica de la fracción derivante de insectos acuáticos en el Rio Vigui, comarca Noggbe-Buglé. Centro Regional Universitario de Veraguas, Facultad de Ciencias Naturales Exactas y Tecnología. Tesis de Grado. Biología. Panamá
Hammer, Ø.; Harper, D. A. T. y Ryan, P. D. (2001). PAST: Paleontological Statistics Software package for education and data analysis. Palaeontologia Electronica, vol. 4(1), 1
Koetsier, P. & Bryan, C. F. (1996). Is macroinvertebrate drift a density dependence mechanism of the benthos in the lower Mississippi river? Journal of Freshwater Ecol-ogy 11(1),1-9.
Lombardo, R & Rodríguez, V. (2008). Calidad biológica del agua en la parte media-baja del Río Santa María, provincia de Veraguas, República de Panamá. Tecnociencia. 10(1):
Matthaei, C. D.; Werthmüller, D.; Frutiger, A. (1998). An update on the reliable quan-tification of stream drift. Archiv. Fur Hydrobiologie. 143(1),1–19. DOI: 10.1127/archiv-hydrobiol/143/1998/1
Needham, P. R. (1928). A net for capture of stream drift organisms. Ecology. (9), 339-342.
Novelo-Gutiérrez, R. (1997a). Clave para la separación de familias y géneros de las náyades de
Odonata de México. Parte I. Zygoptera. Dugesiana 4:1-10.
Principe, R. E. & Corigliano, M. C. (2006). Benthic, drifting and marginal macroin-vertebrate assemblages in a lowland river: temporal and spatial variations and size structure. Hydrobiologia (553), 303-317
Pringle, C. & Ramírez, A. (1998). Use of both benthic and drift sampling techniques to assess tropical stream invertebrate communities along an altitudinal gradient, Costa Rica. Freshwater Biology 39: 359-375.
Quiñones, M.L., Ramírez, J. J. & Díaz, A. (1998). Estructura numérica de la comunidad de Macroinvertebrados acuáticos derivadores en la zona de ritral del río Medellín. Actualidades Biológicas 20(69): 75-86.
Ramírez, A. & Pringle, C. M. (1998). Invertebrate drift and benthic community dynamics in a lowland tropical stream, Costa Rica. Hydrobiologia 386: 19-26.
Ramírez, A. & Pringle, C. M. (2001). Spatial and temporal patterns of invertebrate drift in streams draining a Neotropical landscape. Freshwater Biology 46: 47–62.
Ramírez, A. (2010). Capítulo 5: Odonata. Revista Biología Tropical. Vol.
58 (Suppl.4): 97-136.
Ríos, B. (2008). Comunidades de macroinvertebrados en un río altoandino: Importancia del hábitat, dinámica de la deriva, papel de la materia orgánica y relevancia de la ovoposición. Tesis Doctoral, Departamento de Ecología, Universidad de Barcelona. Barcelona. España.
Rodríguez, B., Ospina-Torres, R., Berrío, M., Cepeda, B., Castellano, G., & Valencia, M. (2006). Variación diaria de la deriva de macroinvertebrados acuáticos y de materia orgánica en la cabecera de un río tropical de montaña en el departamento de Nariño, Colombia. Acta Biológica Colombiana 11, suppl.1. 47-53.
Rodríguez, V. & Cárdenas, X. (2017). Estructura trófica de la entomofauna acuática y calidad del agua de los ríos Zaratí y La Chorrera, provincia de Coclé, república de Panamá. Tecnociencia. Vol. 19, N° 1.
Rodríguez, V. & Duarte, C. (2019). Estructura de la comunidad de insectos acuáticos en la deriva y su relación con la estructura bentónica de la comunidad de insectos, en un tramo del río Santa María, provincia de Veraguas, República de Panamá. Visión Antataura. Vol.3, No.1: 1-23
Rodríguez, V. & Mendoza, M. (2003).Entomofauna acuática asociada al río Agué, en el distrito de la Mesa, Veraguas, Panamá. Tecnociencia 5(2): 109-118.
Rodríguez, V. & Otero, N. (2019). Periodicidad en la deriva de insectos acuáticos, en un tramo del río Gatú, Veraguas, Panamá. Revista Colón Ciencia y Tecnología. 1, 1: 19-29.
Rodríguez, V. & Sánchez, N. (2001). Entomofauna acuática asociada al Río Santa Clara en Veraguas, República de Panamá. Tecnociencia 3(2):73-87.
Aparicio, L & Concepción, A. (2007). Estructura numérica y composición de la
fracción derivante
de insectos acuáticos en un tramo
del río San Pedro, Rincón Sucio, Provincia de Veraguas, Republica de Panamá. Centro Regional Universitario de Veraguas, Facultad de
Ciencias Naturales Exactas y Tecnología. Tesis de Grado. Biología. Panamá.
Community structure in neotropical streams: fish feeding guilds,
disturbance and influence of direct versus indirect effects of predators on
their prey. Ph.D. thesis. University of Maryland, College Park, Washington, D.C.
218p.
Gutiérrez-Fonseca, P. E. (2010). Guía ilustrada para el estudio ecológico y taxonómico de los insectos acuáticos del Orden Coleoptera en El Salvador. En: Springer, M. & J.M.
Sermeño Chicas (eds.). Formulación de una guía metodológica estandarizada para determinar la calidad
ambiental de las aguas de los ríos de El Salvador, utilizando insectos
acuáticos. Proyecto Universidad de El Salvador (UES) – Organización de los
Estados Americanos (OEA). Editorial Universitaria UES, San Salvador, El
Salvador. 64 pp.
Novelo-Gutiérrez, R. (1997b). Clave para la determinación de familias y géneros de las náyades de Odonata de México. Parte II. Anisoptera.
Dugesiana 4:31-40.