Resumen: En este trabajo se presenta una síntesis de las investigaciones realizadas en 25 sitios de la costa sur de la Laguna Mar Chiquita desde distintas líneas de análisis: estudios sobre la historia y evolución biológica de las poblaciones, dieta, usos del cuerpo y estilo de vida a partir de distintos bioindicadores, tales como isótopos estables y microrrestos vegetales, tecnología cerámica y prácticas mortuorias. Los cambios y continuidades detectados son situados en un marco cronológico preciso (32 fechados radiocarbónicos, 4525-375 14C años AP), y con referencia al modelo paleoambiental propuesto para el área. Se propone una ocupación humana desde fines del Holoceno medio, con un hiato entre 3800-2500 años AP y una intensificación notable entre 700 y 500 años AP. Los estudios bioantropológicos permiten diferenciar a estas poblaciones de las que ocuparon la región serrana de Córdoba, y proponen cambios en la composición poblacional a partir de 1200 años AP, sugiriendo similitudes con poblaciones del noreste de Patagonia, noreste de región pampeana y región chaco-santiagueña. Se destaca la importancia del consumo de plantas silvestres durante todo el período, incorporando cultígenos de forma complementaria con posterioridad a 1200 años AP. A partir de este momento se sugiere un incremento en los niveles de actividad física, aumento en la prevalencia de ciertas patologías orales y tendencia secular negativa en la talla de los individuos femeninos. Particularmente a partir de 700 años AP, el crecimiento poblacional, sumado al aumento de las tensiones sociales, podría estar indicando un desmejoramiento en la calidad de vida hacia finales del Holoceno tardío.
Palabras clave: Variabilidad biológica, Bioindicadores dentales, Microrestos vegetales, Cambios entesiales y degenerativos, Isotopos estables, Holoceno Medio y Tardío, Córdoba.
Abstract: This paper presents a synthesis of the archaeological and bioanthropological research carried out at 25 archeological sites on the southern coast of Laguna Mar Chiquita: studies on the history and biological evolution of populations, their diet, body uses and lifestyles, based on diverse bioindicators such as stable isotopes and vegetable micro remains, ceramic technology and mortuary practices. Changes and continuities are determined in a precise time frame (32 radiocarbon dates, 4525-375 14C years BP), and with reference to the paleoenvironmental model proposed for the area. Human occupation is suggested from the end of the middle Holocene, with a gap between 3800-2500 years BP, and notable intensification between 700 and 500 years BP. Bioanthropological studies allow us to differentiate these populations from those in the mountainous region of Córdoba, suggesting changes in their biological composition from 1200 BP and similarities with northeast Patagonia, northeast Pampa and Chaco-Santiago regions. We highlight the importance of wild plant consumption throughout the period, with incorporation of crops after 1200 years BP as a complementary strategy. From this moment on, physical activity levels increase, as well as increased prevalence of certain oral pathologies and a negative secular trend in female height. In particular, from 700 years BP onwards, population growth, combined with an increase in social tension, could indicate a decline in the quality of life towards the end of the late Holocene.
Keywords: Biological variability, Dental bioindicators, Vegetable microremains, Entheseal and degenerative changes: Stable isotopes, Middle and Late Holocene, Córdoba.
Resumo: Este trabalho apresenta uma síntese das pesquisas realizadas em 25 sítios da costa sul da Laguna Mar Chiquita a partir de diferentes linhas de análise: estudos sobre a história e evolução biológica das populações, dieta, usos do corpo e estilo de vida a partir de diferentes bioindicadores, tais como isótopos estáveis e micro-restos vegetais, tecnologia cerâmica e práticas mortuárias. As mudanças e continuidades detectadas estão situadas em um contexto cronológico preciso (32 datações de radiocarbono, 4525-375 14C anos AP), e com referência ao modelo paleoambiental proposto para a área. Propõe-se uma ocupação humana a partir do Holoceno Médio tardio, com um hiato entre 3800-2500 anos AP, com uma intensificação notável entre 700 e 500 anos AP. Os estudos bioantropológicos permitem diferenciar essas populações daquelas que ocuparam a região serrana de Córdoba, e propõem mudanças na composição populacional a partir de 1200 anos AP, sugerindo semelhanças com populações do nordeste da Patagônia, nordeste da região pampeana e região do Chaco de Santiago. Ressalta-se a importância do consumo de plantas silvestres durante todo o período, incorporando cultígenos de forma complementar depois de 1200 anos AP. A partir desse momento, sugere-se um aumento nos níveis de atividade física, aumento na prevalência de certas patologias orais e tendência secular negativa na estatura dos indivíduos do sexo feminino. Particularmente a partir de 700 anos AP, o crescimento populacional, somado ao aumento das tensões sociais, poderia estar indicando uma deterioração na qualidade de vida ao final do Holoceno tardio.
Palavras-chave: Variabilidade biológica, Bioindicadores dentais, Micro-restos vegetais, mudanças entesiais e degenerativas, Isótopos estáveis, Holoceno Médio e Tardio, Córdoba.
Dossier
Vivir en los límites del Gran Chaco y la región pampeana: aportes al estudio de las poblaciones de la costa sur de Laguna Mar Chiquita (Córdoba) desde la Arqueología y la Bioantropología
Living in the borders of the Gran Chaco and the Pampas region: archaeological and bioanthropological contributions to the study of the populations in the southern coast of Laguna Mar Chiquita (Córdoba)
Viver nos limites do Grande Chaco e da região pampeana: contribuições ao estudo das populações da costa sul da Laguna Mar Chiquita (Córdoba) a partir da Arqueologia e Bioantropologia
Recepción: 01 Mayo 2020
Aprobación: 01 Julio 2020
Publicación: 01 Diciembre 2020
Las investigaciones arqueológicas durante las últimas décadas han generado notables avances en torno al conocimiento sobre las poblaciones que habitaron distintos ambientes de la provincia de Córdoba, principalmente la región serrana y los valles adyacentes. Dichas investigaciones permitieron confirmar ocupaciones humanas desde el Pleistoceno final (Rivero, 2007a; Cattáneo et al., 2016; Rivero et al., 2018, 2019), proponer modelos desde el perspectivismo sudamericano (Laguens & Alberti, 2019) y paleoecológicos multi-proxy con base local (Izeta et al., 2017); cambios en las estrategias adaptativas a lo largo del Holoceno (Costa, 2016; Rivero & Medina, 2016), diferencias en los usos del espacio en valles y ambientes serranos (Pastor et al., 2017) y cuestiones vinculadas a procesos identitarios tanto comunitarios como individuales para las comunidades cazadoras-recolectoras y aldeanas (Rivero, 2007b; Fabra et al., 2009; Pastor & Tissera, 2019). La problemática acerca de las prácticas hortícolas y su importancia (relativa) en la alimentación ha sido abordado y discutido desde distintas líneas de evidencia (Laguens, 1999; Laguens et al., 2009; Pastor et al., 2013; Recalde et al., 2017; Fabra & González, 2019).
Sin embargo, la información disponible sobre las poblaciones que habitaron las llanuras del este y noreste asociadas a ríos y lagunas es fragmentada o inexistente. Solo algunas publicaciones entre las décadas 1930-1940 (Aparicio, 1931, 1942; Frenguelli, 1932, 1933) o la mención sobre comunidades Sanavironas por Serrano (1945), que habrían habitado el noreste cordobés y la provincia de Santiago del Estero unos siglos previos a la llegada de los españoles, en vínculo con las poblaciones de las llanuras chaco-santiagueñas en virtud del modelado de vasijas sobre cestos y redes, son las pocas referencias sobre la arqueología de estos grupos humanos. Un trabajo pionero de Montes (1960) postuló un poblamiento de finales del Pleistoceno para la región. A partir de la década de 1990 y primera década del siglo XXI se inician investigaciones arqueológicas orientadas al estudio de la tecnología cerámica de distintos sitios costeros (Bonofiglio, 2004, 2009, 2011). En estas investigaciones, si bien se destacaba la manufactura local, ciertos atributos estilísticos permitían suponer vínculos con poblaciones vinculadas a ríos y lagunas del norte de la región pampeana. En paralelo, y derivado del hallazgo fortuito de restos humanos a partir de 2003, producto de las fluctuaciones hídricas de la laguna (Fabra et al., 2008), se inician estudios bioantropológicos con el objetivo de indagar acerca de la historia biológica de estas poblaciones, en un contexto más amplio y vinculado al poblamiento del centro de Argentina (Fabra et al., 2005, 2007; Fabra & Demarchi, 2009, 2013). Posteriormente, interesó estudiar los cambios y continuidades en los modos de vida, comparando los desarrollos en la región serrana y las comunidades de la costa sur de la Laguna Mar Chiquita, particularmente la alimentación, los recursos vegetales consumidos, los usos del cuerpo y las prácticas mortuorias, desde perspectivas poblacionales (Fabra et al., 2012b, 2014) y recuperando historias de vida en clave osteobiográfica (Canova et al., 2020). Nuevamente, las relaciones establecidas a nivel extraregional particularmente con poblaciones del noreste de las regiones pampeana, patagónica y chaco-santiagueña suponen vínculos biológicos y culturales de larga data (Fabra & Demarchi, 2009, 2013; Nores et al., 2017).
En este contexto, el objetivo de este trabajo es sintetizar los principales aportes realizados desde distintas líneas de investigación arqueológica y bioantropológica durante los últimos 20 años en la costa sur de la Laguna Mar Chiquita. Interesa sistematizar la información y proponer un modelo que permita pensar la dinámica del poblamiento humano durante los últimos 5000 años y discutir vínculos con poblaciones chaqueñas, pampeanas y patagónicas.
La Laguna Mar Chiquita está ubicada entre 26°-32°S y 62°-66°W, en la llanura Chaco-Pampeana, en el centro de Argentina (Piovano et al., 2006), con una extensión aproximada de 600.000 hectáreas (PROMAR, 2015). Este gran humedal abarca la región comprendida por el Distrito Chaqueño Occidental, conformando, con sus áreas circundantes, el extremo sur del Gran Chaco Sudamericano. También se la ubica dentro de la subregión Lagunas Salobres de la Pampa interior (Romano & Brandolin, 2017). Está relacionada con la cuenca Paraná-Plata, y ocupa una depresión tectónica conocida como Pampa hundida cuyo origen se vincula a la falla Tostado-Selva-Melincué (Castellanos, 1959).
Constituye la frontera final de la mayor cuenca endorreica sudamericana, la del Salí-Dulce (Reati et al., 1997), y es la mayor cuenca de estas características en Argentina. Recibe aportes del río Dulce, que se origina al norte, en los Nevados del Aconquija, y desde el sur y sudoeste, el río Suquía o Primero que desemboca en la laguna del Plata, y Xanaes o Segundo, que nacen en las Sierras Pampeanas de Córdoba. En la desembocadura de estos tres ríos se forman bañados que congregan la mayor biodiversidad de la laguna. Está incluida en la Red Hemisférica de Reservas para Aves Playeras (1991), ha sido declarada Reserva de Usos Múltiples por parte de la provincia de Córdoba (Decreto 3215/94), miembro de la Red de Reservas Hemisféricas de Aves Playeras (1991), sitio RAMSAR (2002) y desde 2017 se está trabajando para avanzar en la formalización de un proyecto que declare a este humedal Parque Nacional Ansenuza. Por su extensión constituye la mayor superficie lacustre de Argentina, el mayor lago salado de Sudamérica y el quinto en el mundo.
Esta región ocupa un ambiente transicional entre las provincias fitogeográficas del Espinal y Chaco (Luti et al., 1979). Presenta particularidades que lo definen como un ambiente lacustre. Se lo puede definir por áreas inundables de pastizales y praderas, las cuales por las características salobres del agua permiten el desarrollo de especies halohidromórficas, y en áreas elevadas, bosques aislados (Bucher & Bucher, 2006). En la actualidad, el clima se caracteriza por precipitaciones estivales e inviernos secos. Se cuenta con un sólido modelo paleoclimático para la región basado en el análisis integrado de la información sedimentológica, geoquímica, conjuntos de diatomeas, pigmentos y biomarcadores, con información radiocarbónica sobre restos órganicos (Piovano et al., 2009). Este modelo ha permitido plantear una alternancia entre fases secas y frías, y húmedas y cálidas durante los últimos 13.000 años. A inicios del Holoceno, se habría dado una fase húmeda y cálida, con un balance hidrológico positivo, que fue seguida por una extremadamente seca de la cual no se posee información cronológica del inicio, pero sí de su extensión máxima, que se habría dado hace 4700 años AP. Estas condiciones secas, con hiatus sedimentarios debido a la evaporación, fueron predominantes desde el Holoceno medio hasta mediados del primer milenio, cuando se pueden inferir fases más húmedas hacia 1500 años AP (372–612calAD) y 1100 años AP (920–960calAD). A partir de ese momento, se da un reverso hacia fases secas y frías que no han podido ser datadas debido a hiatus sedimentarios por evaporación (Piovano et al., 2009).
El área de estudio abarcada en el presente trabajo está ubicada en la cuenca baja de los ríos Suquía y Xanaes y áreas adyacentes, en la costa sur de la Laguna Mar Chiquita. Comprende la región ubicada entre 63°03´-62°27´W y 30°34´-30°58´S. (Figura 1). Desde el año 2005 se han prospectado y/o excavado 25 sitios arqueológicos, caracterizados por la aparición en superficie particularmente de restos humanos (Figura 2A, B, C, D, E), hallados de manera fortuita, así como dispersiones de conjuntos cerámicos, materiales líticos o estructuras circulares conocidas como hornitos (Figura 2F). En dichos trabajos, se han recuperado 3 individuos juveniles y 44 adultos (12 femeninos, 27 masculinos, 8 de sexo indeterminado). Respecto a las estructuras circulares, se han relevado 67 asociadas o en cercanías de estructuras de inhumación. Pero debe tomarse a este número como un mínimo, ya que no se ha efectuado una prospección exhaustiva de la costa para identificar concentraciones. Cabe destacar que los museos locales poseen colecciones de estos y otros sitios, recuperados por aficionados y voluntarios de museos, algunos de las cuales han sido incorporadas a esta presentación. En la Tabla 1 se presenta información resumida sobre los sitios arqueológicos, cronología, cantidad de restos humanos asociados, modalidad de inhumación y asociaciones contextuales para los casos que se tiene información.

Figura 1.
Ubicación de los sitios arqueológicos analizados en este trabajo: 1. Miramar, 2. Estancia La Elisa, 3/19/20. La Para (aprox.), 4. Costa Este Laguna del Plata (aprox), 5. Costa oeste Laguna del Plata (aprox), 6. El Diquecito, 7. El Mistolar/Rolo de Depetri, 8. Colonia Muller, 9. Laguna del Plata (aprox), 10. Marull (aprox.), 11. Orihuela, 12. Isla Tigre (aprox), 13. Isla Orihuela, 14. Punta del Silencio, 15. Loma Alta, 17. Playa Grande, 18. Constantino, 21. Campo Bocassi-Agua Mansa, 22. Campo Milesi, 23. Tomasini-La Cerca, 24. Isla Larga, 25. La Loma/Campo Mare

Figura 2.
A. Sitio El Diquecito, Enero 2008; B. conjuntos de material cerámico en superficie, sitio El Diquecito, Abril 2008; C. Excavación entierro cuadricula Q, sitio El Diquecito Abril 2010; D. Excavación, sitio Isla Orihuela, Abril 2011; E. Excavación entierro 4, sitio Punta del Silencio, Agosto 2018; F. Prospección, estructura circular, sitio Rolo de Depetri, Septiembre 2018
Hasta el momento, se han realizado 32 fechados radiocarbónicos sobre restos óseos, 29 mediante espectrometría de masa y 3 por espectrometría de centelleo líquido, los cuales permiten definir un rango temporal para la ocupación humana en la costa sur de Mar Chiquita entre 3240cal AC-1523cal DC (Tabla 2, Figura 3). De estos fechados, se observan tres claras agrupaciones: 4 fechados entre 3240 y 2196 años calAC, 10 fechados entre 816 años calAC y 534 años calDC, y 18 fechados entre 633 y 1523 años calDC. Esta información permitiría inferir una ocupación desde finales del Holoceno medio (agrupación 1), sostenida en el Holoceno tardío[1] desde 2500 años AP (agrupación 2), con una intensificación notable entre 700 y 500 años AP (agrupación 3) (Figura 4). En los tres grupos, y particularmente el primero, los sitios están definidos por hallazgos fortuitos de restos humanos, sin asociaciones contextuales confirmadas. En el grupo 2 se ubicarían las ocupaciones más tempranas del sitio El Diquecito, del cual se recuperaron además de restos humanos fragmentos cerámicos en superficie, y se registraron 32 estructuras circulares de tierra cocida. Finalmente, en el grupo más tardío se destaca el sitio Isla Orihuela costa sudeste, con asociaciones en estratigrafía de restos faunísticos, malacológicos y cerámicos.
Sitios arqueológicos prospectados y/o excavados en la costa sur de la Laguna Mar Chiquita, cantidad de restos humanos y materialidades asociadas. No incluye colecciones de museos regionales

Fechados radiocarbónicos obtenidos en la costa sur de la Laguna Mar Chiquita
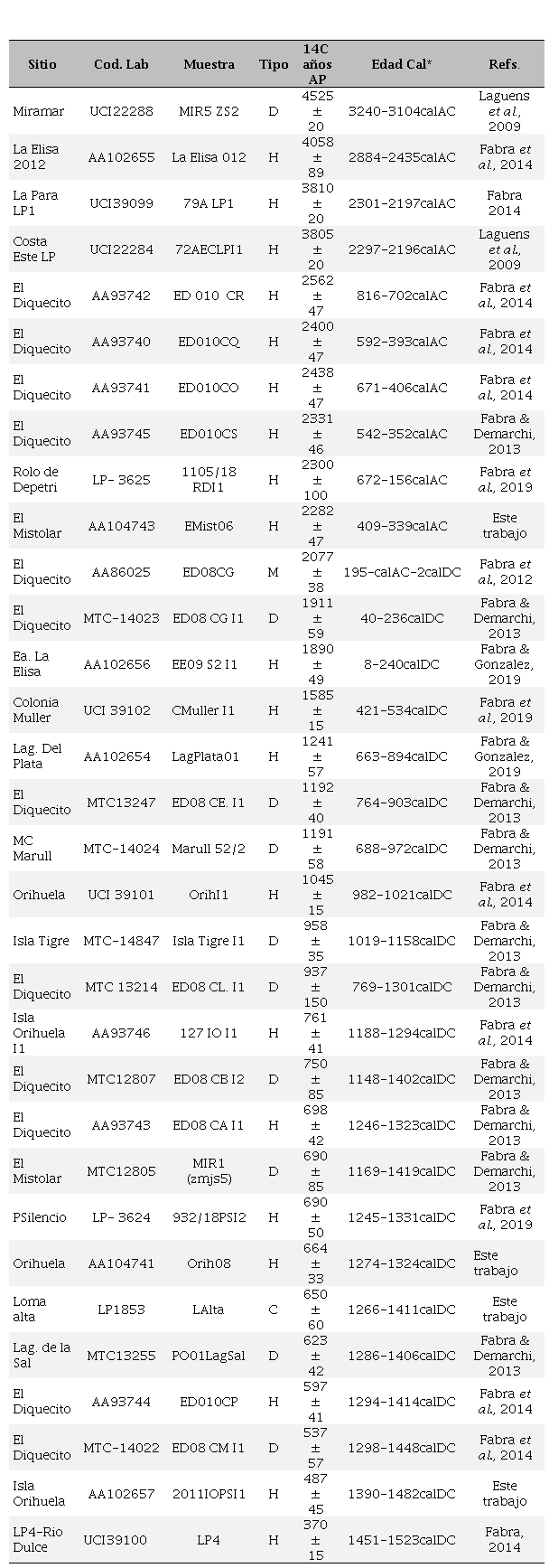
Refs.: Codigo de Laboratorio: MTC (Escuela de Ciencias de Frontera, Universidad de Tokio, Japon), UCI (Universidad de Irving, Estados Unidos), AA (Universidad de Arizona, Estados Unidos), (Laboratorio de Tritio y Radiocarbono, Universdidad Nacional de La Plata, Argentina); Tipo: H: hueso humano, D: diente humano, M: malacologico; C: carbon. * Las calibraciones fueron realizadas con el programa OXCAL V4 3.2 (Ramsey 2017), IntCal 13 curva atmosferica (Reimer et al., 2013)

Figura 3.
Fechados radiocarbónicos calibrados para la costa sur de laguna Mar Chiquita

Figura 4.
Cantidad de fechados radiocarbónicos –sin calibrar–, considerando agrupaciones de 200 años
Esta línea de análisis ha brindado novedosa información, que ha permitido caracterizar la dinámica poblacional desde el estudio de la variación morfológica craneofacial, los rasgos no métricos o discretos del cráneo, y el ADN antiguo. La disponibilidad de información radiocarbónica absoluta permitió situar los patrones de variación observado en un contexto cronológico preciso.
En un primer trabajo (Fabra et al., 2005) desde el análisis de los rasgos no métricos craneales, se indagó acerca de si la variabilidad observada en el registro arqueológico de las poblaciones asentadas en las regiones serrana y llanuras extraserranas tenía su correlato en la composición biológica. A partir de técnicas estadísticas (cluster análisis, autocorrelación espacial, análisis de permutación de respuesta múltiple) se confirmaron las tendencias observadas en el registro arqueológico; posteriormente serían confirmadas también desde el estudio de las prácticas inhumatorias (Fabra et al., 2009), diferenciando a las poblaciones serranas de aquellas que ocuparon la costa sur de Mar Chiquita. También, el análisis de rasgos no métricos permitió sugerir una clara diferenciación de la población de Córdoba (considerando en su conjunto las poblaciones de las sierras y las llanuras) de otras poblaciones de Argentina, aunque se observaron similitudes con poblaciones patagónicas[2]. Posteriormente, utilizando las mismas variables pero otras poblaciones, se profundizó el estudio de la variabilidad en estos rasgos de herencia epigenética a una escala macroregional, considerando el norte de la región pampeana. Se aplicaron técnicas estadísticas de naturaleza exploratoria (cluster analysis, análisis de coordenadas y de componentes principales) y confirmatoria (procedimiento de permutación de respuesta múltiple). Los resultados permitieron inferir una compleja historia poblacional para el área: para las muestras del Humedal del Paraná Inferior se establecieron estrechas similitudes con poblaciones de las sierras y llanuras cordobesas y región chaco-santiagueña, sugiriendo origen común o flujo génico entre esas poblaciones. Para las poblaciones asentadas en el actual territorio uruguayo, se mostraron similitudes con el sur de Cuyo, sur de las sierras pampeanas y centro de Patagonia, no así con el Delta del Parana (Fabra, 2013). En un trabajo reciente en donde sólo se analizaron individuos de la costa sur de la laguna Mar Chiquita, Fabra y colaboradores (2014) estimaron, a partir de análisis de conglomerados y de correspondencia, similitudes entre estas muestras y otras de la región serrana de Córdoba (CBA), región chaco-santiagueña (SGO), así como del Humedal del Paraná Inferior (HPI) y noreste de Patagonia.
Los estudios de morfología craneofacial a partir de morfometría geométrica, con métodos de superposición y deformación (Fabra & Demarchi, 2013), corroboraron las diferencias regionales entre las poblaciones asentadas en las sierras y las llanuras de Córdoba, en distintos momentos del Holoceno, y sugirieron vínculos con el noreste de la región pampeana, región chaqueña y centro (noreste de Chubut) y noreste de Patagonia (desembocadura de Río Negro, isla Gama y San Blas) desde el Holoceno Medio. Estas diferencias se manifestaron a partir de otros indicadores, tales como variables métricas lineales, las cuales permitieron reafirmar vínculos con poblaciones del noreste de la región pampeana y noreste de Patagonia (Fabra, 2008). Para el Holoceno medio, la muestra de Córdoba se ubica a valores cercanos de las muestras del Holoceno tardío de Chaco, del noroeste y noreste de la región pampeana, y muestras tardías del noreste de Patagonia. Para las muestras del Holoceno tardío final, considerando todas las muestras de la provincia de Córdoba, las semejanzas son mayores con el noreste de Patagonia. Los cambios en la morfología craneal son más evidentes en el proceso frontal, más ancho y pronunciado, así como una mayor superficie ocupada por las órbitas y un ensanchamiento en el hueso cigomático y área nasal en las muestras más antiguas respecto a las más tardías.
Estos resultados son similares a los obtenidos por Nores & Demarchi (2011) y, posteriormente, Nores y colaboradores (2011) a partir de análisis de ADN antiguo, quienes estimaron diferencias estadísticamente significativas entre los habitantes de las sierras y de la llanura de Córdoba a partir de 1200 años AP. Estas diferencias se manifiestan particularmente en la mayor prevalencia del haplogrupo B en las sierras y la mayor prevalencia de los haplogrupos C y D en llanura. Se sugiere un aporte inmigratorio diferente en ambas regiones, a partir de 1200 años AP, con poblaciones de Patagonia para las poblaciones asentadas en Mar Chiquita. En un trabajo de 2014, Fabra y colaboradores estimaron a partir de un mayor número de muestras de la costa sur de Mar Chiquita (41 analizadas, 38 tipificadas) la alta frecuencia del haplogrupo C (55%), mientras que los haplogrupos A y D presentan prevalencias intermedias (16% y 26%, respectivamente) y el haplogrupo B está apenas representado por un único individuo.
Más recientemente, se analizó la región hipervariable I del ADN mitocondrial (RHV-I) de 11 individuos del sitio El Diquecito (costa sur Mar Chiquita), 4 corresponden al período pre-1200 AP y 7 a la etapa post-1200 AP (Nores et al., 2017). La totalidad de los individuos previos a 1200 años AP tienen haplogrupo C, mientras que en las muestras posteriores a 1200 años AP 3 individuos tienen el haplogrupo C y 4 el haplogrupo D. Si bien es un número muy reducido de individuos, estarían sugiriendo cambios en la composición poblacional en ambos momentos, como se sostuvo en trabajos previos, con otros marcadores (Fabra & Demarchi, 2013). El análisis de la secuencia de la región control del ADN mitocondrial permitió recuperar 7 secuencias correspondientes al haplogrupo C1 –con dos haplotipos diferentes, uno nodal– y 3 al D1 –con 3 haplotipos diferentes, D1g, D1j y nodal–. Respecto a la procedencia geográfica, uno de los linajes identificados –C1+16092C+16390– no había sido descripto con anterioridad, por lo cual podría tratarse de un linaje extinto o en baja frecuencia en la población actual (Nores et al., 2017). Por otra parte, el linaje D1g está altamente representado en poblaciones antiguas y actuales patagónicas, y actuales de las provincias de Córdoba, San Luis y Santiago del Estero (Pauro et al., 2013). Su presencia en muestras tardías de la costa sur de Mar Chiquita nos haría pensar en contactos recientes con poblaciones del sur del país. Es de interés la continuidad en el tiempo que se puede establecer con la población que actualmente vive en la localidad de La Para, cercana al sitio El Diquecito en 15 km, dada por la alta frecuencia de los linajes C, frecuencias similares para A y D y la ausencia o muy baja frecuencia del haplogrupo B en ambas (García & Demarchi, 2009; Fabra et al., 2014). Finalmente, los cambios que se dieron alrededor de 1200 años AP en la composición biológica de la población asentada en la costa sur de Mar Chiquita podrían explicar, parcialmente, la tendencia secular negativa observada en la talla de los individuos femeninos, no así en los masculinos[3](Loupias & Fabra, 2019).
Las primeras referencias a las ocupaciones humanas en el área, y particularmente, la caracterización de materiales cerámicos, corresponden a los trabajos de Frenguelli (1932), Aparicio (1942), Serrano (1945) y Oliva (1947), quienes destacan la presencia de impresiones de redes y cestería, así como incisiones, sugiriendo semejanzas con las sierras de Córdoba, región chaco-santiagueña y el litoral argentino.
Si bien hasta el momento no ha habido investigaciones orientadas a conocer las fuentes de aprovisionamiento de rocas y minerales, entre ellos arcillas, así como estudios de tecnología lítica de las colecciones de los museos locales, o de los materiales recuperados en excavaciones sistemáticas, se pueden mencionar algunas consideraciones generales sobre estas tecnologías.
En el caso de los artefactos líticos, Bonofiglio (2004, 2016) menciona el hallazgo de puntas de tipo lanceoladas, de entre 5 y 8 cm., confeccionadas en cuarzos, cuarcitas y calcedonias, así como triangulares (Figura 5b) recuperadas en las playas de Laguna del Plata, Campo Mare, las barrancas del sitio Loma Alta o Playa Grande, así como un gancho de propulsor de piedra en el sitio Estancia La Elisa. Si bien no tienen cronología absoluta asociada, por sus similitudes morfológicas con artefactos recuperados en las Sierras Centrales los adscribe a grupos cazadores-recolectores del Holoceno Temprano y Medio (González, 1960; Rivero & Heider, 2017; Pautassi & Sario, 2018). Son poco frecuentes los hallazgos de piedras grabadas (Figura 5a). También menciona el hallazgo de artefactos como morteros y sus manos, conanas, cuchillos (Figura 5c), sierras, bolas con y sin surco, raspadores y hachas (Bonofiglio 2004, 2016).
En cuanto a la cerámica, para el sitio Loma Alta, fechado en 650 ± 50 14C años AP, describe una gran cantidad de formas (Bonofiglio, 2010), particularmente ollas (Figura 5I, L) y pucos o escudillas (Figura 5J). Otro tipo de contenedores identificados han sido las denominadas “botellas”, de forma restringida independiente, o vasos, de forma restringida simple. Se han recuperado contenedores de gran tamaño, definidos como platos grandes (op. cit., p. 14), redondeados de paredes divergentes y bases planas o plano-cóncavas. En el caso de estos contenedores, no tienen evidencias de haber sido sometidos al fuego, para la cocción de alimentos, más bien hace suponer que se usaron para servir alimentos a varias personas, en celebraciones comunitarias cotidianas o especiales. Se han identificado contenedores clasificados como cucharas, con o sin mango, y un fragmento del extremo distal de una pipa (op. cit., p. 16), que permite suponer el consumo de estimulantes o alucinógenos.
Del total, un 30% presenta huellas de uso asociadas a la exposición al fuego y un 20% impresiones de redes, cestas o su combinación en su interior (Figura 5K) (Bonofiglio, 2010, p. 12). En estos casos, se ha sugerido que la manufactura de las vasijas se realizó a partir de una cesta, cubierta con redes en algunos casos, a los fines de agilizar su manufactura. También, para imprimir el diseño presente en las cestas en estos contenedores. Dado que los fragmentos que presentan estas impresiones carecen de restos de hollin u otras adherencias, se ha propuesto su función asociada a la contención de elementos sólidos, posiblemente frutos (Bonofiglio, 2010, p. 13).
Se reportan hallazgos de fragmentos cerámicos en sitios costeros tales como La Playa (Bonofiglio, 2004), Campo Mare, El Camping o Laguna del Plata (Bonofiglio, 2016). La composición de las pastas es de cuarzo y feldespato, arena y tiestos molidos, así como algunos vegetales; es escasa la presencia de mica. Las técnicas de manufactura son a partir de rollos, trabajados en espiral, modelado y moldeado sobre cestas (Bonofiglio, 2009). Los acabados superficiales son por lo general alisados y pulidos. Bonofiglio (2016) define la tecnología cerámica como expeditiva, con escasa selección de materias primas, y cocida en fogones abiertos, lo cual resulta en piezas con cocción irregular. La decoración en las vasijas presenta motivos variados: modelados, zig-zag, cepillados, líneas rectas u ondulantes, rellenas con puntos, que componen guardas simples o complejas (Figura 5G, H), en algunos casos, pintadas de rojo o negro (Bonofiglio, 2004; 2010, p. 14). Es interesante vincular esos motivos con representaciones sobre el agua, en términos de Eliade (2010). También pueden ser vinculados con ambientes acuáticos la existencia de redes y cestas, materializada en las impresiones, o su combinación, en las superficies internas o externas de las vasijas. Este último punto debe abordarse con mayor profundidad, para confirmar si estas impresiones forman parte de decisiones conscientes de los alfareros, para transmitir mensajes a través del entramado, o responde a cuestiones tecnológicas vinculadas con el propio proceso de manufactura sobre este tipo de contenedores. Son frecuentes los modelados o apliques, con representaciones zoomorfas de aves (Figura 5D, E, H), miniaturas (5F) y estatuillas antropomorfas planas (Figura 5M). Se han descripto otros objetos clasificados como torteros, brazaletes y estatuillas, artefactos cónicos o vasos, todos ellos con decoración incisa (Bonofiglio, 2004, p. 35).

Figura 5.
A. Piedra grabada, posible afilador, sitio Loma Alta; B. punta de proyectil triangular apedunculada, sitio Loma Alta; C. raedera, sitio Loma Alta; D. aplique zoomorfo, sitio Loma Alta; E. aplique, representación de búho, sitio Loma Alta; F. Miniatura de vasija, sitio Loma Alta; G. borde con motivos incisos, de tipo surco rítmico punteado, sitio Loma Alta; H. vaso hueco zoomorfo, representación de ave, sitio Loma Alta; I. vasija entera, sitio Loma Alta; J. cuenco reconstruido, con incisiones y borde en ondas, sitio Loma Alta; K. fragmento con impresión de redes y cestas, pared interna, sitio El Diquecito; L. vasija con asa, Laguna del Plata; M. estatuilla antropomorfa, de tipo placa incisa, sitio Loma Alta.
Relacionado con la práctica alfarera, podemos incluir la construcción de estructuras semisubterráneas realizadas con arcilla, denominados comúnmente hornillos o botijas (Figura 2F). Fueron mencionados por primera vez en trabajos de Aparicio (1931) y Frenguelli (1933). Si bien son estructuras de aparición frecuente en la región central de Argentina, incluyendo las provincias de Santa Fe, Santiago del Estero, Norte de Córdoba, La Rioja, San Luis y Mendoza, también se los ha identificado en Jujuy y Norte de Formosa, asociados a sitios cuyas cronologías los ubican en momentos de condiciones más frías y secas que las actuales (Lamenza et al., 2019). En la costa sur de Mar Chiquita se han detectado varios cientos, particularmente en los sitios El Diquecito (N= 32), Punta del Silencio (N= 16), Rolo de Depetri (N=14), Loma Alta (N=6, Giraudo, com. pers), Campo Fauda (11, Giraudo, com. pers.), Isla Tigre (50, Giraudo, com. pers.), e Isla Larga, en este último caso más de 700 hornillos fueron relevados (Giraudo, com. pers). Se debe tener en cuenta que la visibilidad de los mismos está directamente relacionada con los niveles bajos de la laguna, estando muchas estructuras sumergidas en épocas de niveles altos. Respecto a su funcionalidad, se proponen tanto usos vinculados a la cocción de alimentos (Cornero & Ceruti, 2009; Cornero et al., 2013), o al almacenamiento en situaciones de crisis ambiental (Laguens & Bonnin, 2009). En los casos que fueron excavados en la costa sur de Mar Chiquita, además de semillas carbonizadas de Prosopis sp., se recuperaron restos de fauna –Dasypodidae, cascaras de huevo de Rhea americana y Lama guanicoe–, o fragmentos de cerámica (Bonofiglio, 2016; Giraudo, com. pers.). En otros casos, las estructuras fueron parcialmente destruidas para excavar las fosas donde inhumarían sus muertos, como en el caso del individuo de la cuadricula L en el sitio El Diquecito –937 ± 150 años 14C AP[4]– (Fabra et al., 2012b), lo cual hace suponer que el hornillo no se encontraría en uso. En otros casos, en momentos más tardíos, los individuos fueron enterrados a escasos centímetros de los hornillos, como el individuo 2 del sitio Punta del Silencio, fechado en 690 ± 150 años 14C AP (Fabra et al., 2019), o el Diquecito –individuo A, 698 ± 42 años 14C AP–.
Se ha considerado a estas estructuras semisubterráneas como indicadoras precisas de climas áridos o semiáridos, asociados a la región del Espinal (Cornero et al., 2013). Para los casos mencionados, sus cronologías coinciden parcialmente con períodos de menor disponibilidad de agua (Piovano et al., 2009), por lo que podemos suponer se hallaban en funcionamiento. Si consideramos la propuesta de Cornero et al. (2013) y del Rio et al. (2016) de que la cantidad de estructuras podría relacionarse a la movilidad de los grupos humanos, y de Laguens (1999) y Laguens & Bonnin (2009) que los vinculan al almacenamiento como parte de estrategias previsionales en épocas de sequía, podemos pensar en El Diquecito y Punta del Silencio como sitios que fueron poblados de manera reiterada e intensiva, o con sedentarismo ocasional. En un contexto ambiental de crisis hídrica, el almacenamiento de un alto volumen de recursos en lugares de acceso restringido a ciertas comunidades podría ser una estrategia práctica.
Los rituales funerarios son complejos sistemas simbólicos en los cuales el entierro, la ubicación intencional que se les dé en la tumba, es sólo uno –quizás el más visible y que ofrece evidencia material– de numerosos actos simbólicos que preceden y suceden a la práctica inhumatoria (Bartel, 1982). Como tales, permiten recuperar prácticas y decisiones de los vivos, de las comunidades de pertenencia de los difuntos, vinculadas a la muerte, como pocos registros materiales lo permiten. En este contexto, el estudio de las modalidades de entierro fue una de las problemáticas que mayor interés generó. En un primer momento (Fabra et al., 2009), interesó indagar si había un correlato entre las diferencias que varios autores habían detectado en distintos aspectos de la cultura material, en distintas regiones de la provincia para momentos tardíos (Serrano 1945; Laguens & Bonnin, 2009), y las modalidades de inhumación. Como se mencionó previamente, estudios de variabilidad craneofacial y posteriormente de ADN antiguo habían sugerido diferencias en cuanto a la composición biológica entre distintas regiones geográficas de Córdoba. Para el noreste de la provincia, se describió la modalidad de inhumación de 6 individuos, todos con información cronológica por AMS asociada, considerando variables como el sexo, la edad, el tipo de entierro, la posición del esqueleto, la disposición de miembros inferiores y superiores, y la orientación del esqueleto. Se analizó esta información, junto a la obtenida para otros individuos de la provincia, mediante análisis estadísticos (análisis de permutación de respuesta múltiple, análisis de componentes principales, test de chi cuadrado). Los resultados confirmaron la existencia de un patrón de agrupamiento geográfico regional, con diferencias estadísticamente significativas entre las modalidades de entierro practicadas por las distintas comunidades en el Holoceno tardío. Particularmente para la costa sur de Mar Chiquita, las inhumaciones se caracterizaban por entierros primarios simples, decúbito dorsal o ventral (en menor medida), con los miembros inferiores extendidos o flexionados hacia derecha o izquierda. En otras regiones de la provincia, las modalidades de entierro más frecuentes son primario simple decúbito lateral, no habiéndose registrado hasta el momento entierros en posición decúbito ventral y particularmente dorsal con miembros extendidos. También se detectaron en general diferencias estadísticamente significativas por sexos: los individuos masculinos estaban predominantemente orientados con el cráneo al sur y los pies al norte, y las mujeres, con el cráneo al oeste y los pies al este. Si estos rituales permiten expresar las relaciones entre vivos y muertos en una sociedad, podríamos suponer que estas diferencias dan cuenta de las distintas concepciones que tenían las comunidades serranas y las de la llanura frente a la muerte.
Si bien ese trabajo inicial contó con un n muestral reducido (N=19), permitió confirmar las claras diferencias en torno al tratamiento de los muertos que existían, en líneas generales, entre las comunidades serranas y aquellas asentadas en las llanuras, tanto en entornos fluviales como lacustres. Tomando de base ese trabajo inicial, nos preguntamos si las comunidades asentadas en el entorno lacustre de Mar Chiquita habían mantenido esa modalidad de entierro a lo largo del Holoceno tardío sin cambios, o si podían distinguirse ciertos cambios, y en tal caso, a que otros aspectos sociales o culturales podrían asociarse (González & Fabra, 2019). Se confirmó la tendencia sugerida en el trabajo de Fabra y colaboradores (2009) respecto a la predominancia de entierros primarios simples (un único caso de un entierro primario, doble), decúbito dorsal (un único caso decúbito ventral) con brazos en posición extendida o flexionados sobre el tórax. La posición de los miembros inferiores resultó con prevalencias similares para las posiciones extendidos, flexionados hacia la izquierda o hacia la derecha. Estas modalidades están presentes a lo largo del Holoceno; sin embargo, se observan diferencias entre hombres y mujeres en la disposición de los miembros superiores, siendo más frecuente para las mujeres, en momentos previos a 1200 años AP, los miembros extendidos, mientras que para los hombres las posiciones son más variables. En momentos posteriores, las mujeres presentan más variabilidad, similar a los hombres, y en este caso, se abandonan algunas posturas (hiperflexionamiento sobre el tórax). Otra diferencia que interesa destacar, en esos dos momentos del Holoceno tardío, es la orientación del cuerpo para individuos femeninos: en momentos previos a 1200 años AP, los cuerpos de las mujeres se orientaban con la cabeza hacia el este o el oeste, y en momentos posteriores, predomina la posición norte o sur. En el caso de los individuos masculinos, el patrón es inverso, en sentido norte sur (González & Fabra, 2019). Si bien las modalidades predominantes están presentes desde al menos 4000 años AP, los sutiles cambios detectados (sea en la orientación del cuerpo o la ubicación de los miembros superiores e inferiores)– ocurren a partir de 1200 años AP, y pueden estar relacionados a cambios en la organización social de estas comunidades por la incorporación de ciertos cultivos, a pequeña escala, así como vínculos con comunidades asentadas en regiones cercanas. Sobre este punto avanzaremos más adelante.
Las excepciones a estos patrones están dadas por entierros que por sus características pueden considerarse inusuales, sea por los materiales que los acompañan, o las lesiones óseas que presentan los individuos[5]. Ambos casos fueron presentados y discutidos por Canova et al. (2020) y por Fabra et al. (2015) respectivamente. En uno de los casos, el individuo femenino está enterrado decúbito ventral, con las manos y pies hacia atrás, como si hubieran estado atados. Está acompañado por una valva de molusco sin formatizar, de la especie Anodontites trapesialis que provendría de ríos o afluentes como el rio Paraná, Dulce, Carcarañá o Tercero, y que puede considerarse evidencia del contacto con poblaciones de esas regiones (Fabra et al., 2012a, Canova et al., 2020). Otras especies de moluscos han sido reportadas en sitios del Paraná Medio como ajuar funerario (Ramos van Raap, 2018). Otro hallazgo que interesa destacar, por la presencia de moluscos asociados a restos humanos es el del sitio Paraje Buey Muerto, distante a 50 km al sur de Mar Chiquita, sobre la cuenca media del río Suquía (Gordillo & Fabra, 2018). En este caso, se identificaron cuentas discoidales y troncocónicas, que si bien estaban confeccionadas sobre una especie local (Megalobulimus lorentzianus), remiten en su forma y técnica de manufactura, particularmente las troncocónicas –zona umbilical de la concha de un ejemplar de caracol– a otras recuperadas por Lamenza et al. (2016) en el sitio El Quebracho, en el sector Pilcomayo chaqueño (Lamenza et al., 2019).
Estos hallazgos, si bien infrecuentes, invitan a pensar en las relaciones entre humanos y otros animales, vivos y muertos, rituales mortuorios y su relación con identidades sociales, temas que usualmente no son abordados en la arqueología de Córdoba en general, y en particular, en la arqueología de la costa sur de la Laguna Mar Chiquita. Hasta la fecha, sólo se ha reportado el hallazgo de un dasipódido (Chaetophractus sp.) asociado a restos humanos (ca. 2100 años AP) en el Valle de Traslasierra (Pastor et al., 2017). En el sitio Punta del Silencio (690±50 años 14C AP), se recuperaron 12 especímenes de distintos vertebrados, entre ellos macho y hembra de la especie Mazama gouazoubira (corzuela) (Izeta, 2018, ms), asociados a un individuo adulto femenino (individuo 4). Este último fue hallado semisumergido, parcialmente desarticulado en la región pélvica. No se encontró el cráneo, pero el resto del esqueleto postcraneal fue recuperado casi en su totalidad bajo el agua, y por la ubicación de las distintas partes anatómicas, se supone estuvo enterrado en posición decúbito dorsal, cráneo orientado al Norte, pies hacia el sur, con las manos ubicadas al costado del cuerpo (Fabra et al., 2019). Esta posición de entierro coincidiría con el patrón observado para individuos femeninos a partir de 1200 años AP (González & Fabra, 2019). Fabra et al. (2019) han sugerido, como una primera aproximación, la idea de pensar en estos animales incorporados al ritual mortuorio como parte de la identidad social de la persona (Queiroz et al., 2017) o psicopompas, animales guías para el alma del difunto (Goepfer, 2008).
En cuanto al lugar elegido para las inhumaciones, el entierro dentro de los sitios habitacionales, que promueve y mantiene el vínculo entre vivos y muertos, es común en la costa sur de Mar Chiquita, así como en otros sitios del Chaco meridional (Calandra & Salceda, 2006; Desántolo et al., 2005). En sitios como El Diquecito (Fabra et al., 2008), donde coexisten áreas de inhumación con otras evidencias materiales que hacen suponer espacios para actividades múltiples, se daría esa coexistencia entre lo doméstico y lo ritual, lo cotidiano y sagrado, como ha sido propuesto por Ramos van Raap (2018) para sitios del Delta del Paraná.
Hasta el momento, se han identificado dos contextos compatibles con situaciones de violencia interpersonal en los sitios costeros de Estancia La Elisa –EE09– (1890±49 14C años AP) y El Diquecito –ED08– (750±85 14C años AP). Ambos casos fueron discutidos por Fabra et al. (2015) en el marco de la evidencia regional, y se los adscribe, por las características de las lesiones y la ubicación de los proyectiles, tanto a situaciones de enfrentamientos cara a cara –ED08 individuo 2– como a cierta distancia, por el uso de proyectiles con puntas realizadas con material lítico –EE09 individuo 1– u óseo –EE08 individuo 1– y su combinación con boleadoras u otros objetos de punta roma –ED08 individuo 2–. En los tres casos, se encuentran involucrados individuos masculinos. En cuanto a las puntas de proyectil confeccionados sobre hueso, su diseño y decoración es similar a otras encontradas en la región chaco-santiagueña (Laguens & Bonnin, 2009) y en el Litoral, particularmente el Delta del Paraná (Pérez Jimeno & Buc, 2010).
De forma paralela a las líneas de investigación mencionadas previamente, se llevaron adelante estudios bioarqueológicos tendientes a abordar cuestiones vinculadas con la alimentación, relevando distintos biondicadores de salud oral (Fabra et al., 2012b), isótopos estables (Fabra & González, 2019) y microrestos vegetales contenidos en tártaro dental (Tavarone et al., 2019). En línea con los objetivos del proyecto marco general, en estas investigaciones interesó estudiar cambios y continuidades en las estrategias de subsistencia, y el efecto que habría tenido en estas comunidades cazadoras-recolectoras-pescadoras la incorporación del cultivo a pequeña escala. En relación con este último punto, interesaba conocer la cronología de este proceso, y las especies vegetales incorporadas, ya que no había ninguna información hasta el momento para esta región.
Una primera aproximación al estudio de las patologías de origen metabólico e infeccioso/degenerativo es el trabajo de González & Fabra (2011), en el cual analizaron 34 individuos de la costa sur de Mar Chiquita, y sugirieron que la mayor prevalencia de individuos con hipoplasias del esmalte (6,97%), pérdidas dentales antemortem –PDAM– (20,28%) y abscesos (9,49%) hacia finales del Holoceno tardío se podía relacionar con un desmejoramiento en las condiciones de salud. En un trabajo posterior (Fabra et al., 2014), donde se analizaron exclusivamente 26 muestras de la costa sur de la Laguna Mar Chiquita, se detectaron prevalencias similares de hipoplasias del esmalte dental (6,51%), PDAM (17,35%) y abscesos (7,69%), siendo más prevalentes en femeninos las PDAM (29,58%) y en masculinos las hipoplasias del esmalte (10,36%). Para todos los indicadores, caries incluidas, se observa un incremento progresivo con la edad de muerte, con excepción de las hipoplasias del esmalte. Las PDAM (18,09%) y las hipoplasias del esmalte (7,65%) serían más frecuentes en el período posterior a 1200 años AP.
En cuanto al desgaste dental, particularmente los adultos medios y mayores de sexo masculino, han presentado los grados más severos, en comparación con los de la región serrana (González & Fabra, 2018). Las diferencias son significativas también en la comparación cronológica, siendo más severos en momentos tardíos del Holoceno. En este período se observa que el desgaste no se limita solo a molares y premolares, sino que está presente en la dentición anterior. En la costa sur de Mar Chiquita se presenta además la mayor variabilidad en los patrones de desgaste (oblicuo, plano, cóncavo, redondeado) con predominancia de formas planas y cóncavas, resultado de altas presiones masticatorias. Se destacan las formas oblicuas en individuos femeninos y redondeadas en masculinos. Se han detectado surcos, los cuales junto con las formas oblicuas y redondeadas han sido asociados a actividades paramasticatorias. Las autoras concluyen que los cambios en las frecuencias y particularmente en los patrones están más relacionados con el tipo de alimentos –fibrosos y duros–, las técnicas de procesamiento –secado, salado de alimentos, molienda que involucra inclusión de partículas abrasivas a las preparaciones– más que con los recursos consumidos, como lo sugiere también el registro arqueobotánico (Tavarone et al., 2019) y los datos isotópicos (Fabra & González, 2019).
En un trabajo sobre 41 individuos (Fabra et al., 2012b), los indicadores de salud oral mostraron prevalencias bajas de caries (11,52%) y elevadas de cálculo dental (61,93%); en el caso de las caries, más prevalentes en mujeres (12,14%), con un leve aumento hacia finales del Holoceno tardío. Un estudio posterior, con un n muestral menor –26 individuos– (Fabra et al., 2014), permitió detectar prevalencias similares para caries (13,03%), mayor en individuos femeninos (15,26%). Los valores levemente mayores en femeninos no estarían indicando diferencias en cuanto a los alimentos consumidos o hábitos de higiene, sino que se relacionarían con cambios hormonales. Las caries serían más frecuentes en momentos previos a 1200 años AP (14,28%), aunque este valor puede estar subrepresentado por las pérdidas dentales antemortem. Nuevamente, estas pérdidas son mayores en individuos femeninos. En conjunto, estos valores son coincidentes con estrategias basadas en la caza y recolección, o incorporación en menor grado de cultivos a la dieta (Fabra & González, 2019). Los valores similares de frecuencias tanto en momentos iniciales y finales del Holoceno estarían sugiriendo el consumo de alimentos potencialmente cariogénicos, es decir ricos en azucares y/o carbohidratos.
Los análisis de microrestos vegetales contenidos en tártaro dental en individuos recuperados del sitio El Diquecito permitieron confirmar el consumo y/o manipulación de Geoffroea decorticans, Prosopis sp., y Trithrinax campestris al menos desde 2500 años AP con una incorporación paulatina de Phaseolus sp. alrededor de 1200 años AP, seguida de Cucurbita sp. y Zea mays hace 1000 años AP (Tavarone et al., 2019). El consumo de vegetales silvestres, entre ellos los frutos de palmeras, así como el complejo maíz-poroto-zapallo producido a pequeña escala ha caracterizado la subsistencia de poblaciones asociadas a la entidad arqueológica Goya-Malabrigo (Bonomo et al., 2011a; Bonomo, 2012; Politis & Bonomo, 2012; 2018; Colobig et al., 2018). La cronología de la incorporación de frutos de palma, maíz, poroto y zapallo en la costa sur de Mar Chiquita sería posterior a los procesos ocurridos en el Delta del Paraná, en poblaciones asociadas a la entidad arqueológica Goya Malabrigo (Politis & Bonomo, 2012, 2018, Colobig et al., 2015; Cornero & Rangone, 2015). En la región serrana de Córdoba, las evidencias serían más tempranas, particularmente para el maíz, a inicios del Holoceno tardío –ca. 3000 años AP– (Pastor et al., 2012).
Las tendencias sugeridas por los bioindicadores orales, y la evidencia arqueobotánica se complementan con los datos isotópicos para la región: para momentos previos a 1200 años AP, los valores de δ13C sobre colágeno tienen un rango entre -13,7‰ y -17,3‰, con un promedio de -15,5 ± 1,8‰ (n = 10), y de 9,4‰ para la única muestra de δ15N. Los valores son similares en momentos posteriores, entre -11,8‰ y -17‰, con un promedio de -15,0 ± 1,5‰ (n = 17), y para nitrógeno, 7,9‰ a 12,9‰, con un promedio de 10,4 ± 1,7‰ (n = 10) (Fabra & González, 2019). Estos valores no sugieren cambios de importancia en la alimentación, y confirman que el cultivo fue complementario a las estrategias cazadoras-recolectoras en esta área, de manera similar a los procesos ocurridos en las Sierras Centrales (Laguens, 1999; Medina et al., 2014, 2016) o Delta del Paraná Inferior (Bonomo et al., 2011b). Los valores elevados de δ13Ccol y enriquecidos de δ15N sugieren consumo principalmente de vegetales con fotosíntesis C3 –entre ellos zapallo y poroto– o herbívoros consumidores de dichas plantas, animales adaptados a ambientes lacustres, y roedores.
Se relevaron osteofitosis y nódulos de Schmorl para reconstruir los niveles de actividad física. Se detectaron lesiones en codos (10%), tobillos y pies (8%) y el segmento vertebral L1-L5 (78%), a lo que se agrega una mayor prevalencia de lesiones en las vértebras cervicales (48%) y torácicas (70%), particularmente en adultos jóvenes[6]. Se identificaron nódulos de Schmorl en la porción lumbar de individuos jóvenes masculinos, a finales del Holoceno tardío. Los individuos de esta región presentan mayor afectación que los de las sierras de Córdoba, con mayor cantidad de porciones anatómicas comprometidas, en columna vertebral –segmentos bajos- y esqueleto apendicular, particularmente rodillas y pies (Fabra et al., 2012b).
En un trabajo posterior, con un menor número de muestras (N=9) pero de procedencia exclusiva de la costa sur de la Laguna Mar Chiquita (Fabra et al., 2014), se detectó un aumento en la prevalencia de osteofitosis en adultos jóvenes (alcanzando el 100% en la porción T1-T6 y L1-L5) y de nódulos de Schmorl en adultos jóvenes y medios, hacia fines del Holoceno tardío (20% en ambos). En cuanto a articulaciones del esqueleto apendicular, si bien en momentos tardíos disminuyen levemente las prevalencias, aumenta la cantidad de porciones afectadas, apareciendo lesiones en tobillos y pies, en adultos jóvenes. Se relevó la eburnación[7] como otro indicador de cambio degenerativo en las articulaciones: estuvo presente en ambos períodos, afectando distintas articulaciones, siendo mayor en adultos medios, en la muñeca (41,7%) en el primer período y el hombro (30%) para el segundo. La eburnación en articulaciones de miembros superiores hace pensar en un mayor uso de esas porciones anatómicas, en una amplia variedad de actividades. Las lesiones en adultos jóvenes han sido interpretadas como debido a un inicio temprano de actividades exigentes desde el punto de vista físico y al mantenimiento prolongado de posturas en cuclillas o arrodilladas (Salega & Fabra, 2013). La presencia de nódulos de Schmorl sugiere actividades de carga o traslado de objetos pesados, o actividades repetitivas que requieren cierto esfuerzo (Arrieta & Mendonca, 2011; Salega & Fabra, 2017). El aumento de afectación de las lesiones degenerativas en el esqueleto apendicular para los momentos finales del Holoceno puede estar vinculado al mayor esfuerzo físico derivado de nuevas actividades, o una acentuación de las que ya se venían realizando, producto de una intensificación económica, no vinculada exclusivamente con la práctica hortícola, sino también con las condiciones ambientales, el aumento poblacional y potenciales conflictos asociados (Fabra et al., 2012b; Salega & Fabra, 2013).
Las investigaciones realizadas hasta el momento permiten sugerir que circa 4500 antes del presente se habían establecido en las costas de la laguna Mar Chiquita poblaciones con economías cazadoras-recolectoras, en un contexto ambiental caracterizado por una de las más severas fases hidrológicas negativas que ocurrieron en la región, cuyo máximo ocurrió hace 4700 años AP (Piovano et al., 2009). El registro arqueológico de inicios del Holoceno tardío está representado exclusivamente por restos humanos recuperados en los sitios Miramar (Mir5), Estancia La Elisa (EE12), La Para (LP1, LP) y Costa este de Laguna del Plata (CELP). A partir de 3800 años AP la señal arqueológica desaparece, y vuelve a haber indicios de ocupación humana recién 2500 años AP. Si bien las cronologías de las épocas de sequía no han podido resolverse por la aparición de hiatos sedimentarios indicados por capas de yeso-halita (Piovano et al., 2009), es sugerente pensar que la falta de indicios arqueológicos de ocupaciones humanas pueda deberse, más que a un sesgo arqueológico, a una real desocupación del área por al menos 1000 años, si consideramos el volumen de datos radiocarbónicos disponibles (ver Figura 4). Estas cronologías coinciden con la reconstrucción de los niveles de paleolago realizada por Piovano et al. (2009), donde se evidencia una gran variación hidrológica, predominando fases de alta evaporación –algunas extremas– desde 4500 años AP, hasta al menos 2000 años AP. Esto habría originado una disminución en el caudal de los ríos, transformando en pantano la laguna, y generado una merma en la fauna y la flora disponibles en este ambiente. Seguramente estas poblaciones se organizaron como bandas, que cazaron y recolectaron recursos naturales del bosque chaqueño, tales como guanacos, venados de las pampas, ñandús, pecarís, y frutos como el algarrobo, chañar y mistol[8].
Los cráneos fechados a inicios del Holoceno tardío presentan una deformación artificial, de tipo circular, que luego no vuelve a registrarse en el área, en momentos tardíos. De estos momentos es el registro del haplotipo D1j. Su representación en poblaciones del centro oeste argentino es elevada –45%– y fue identificado en el individuo del sitio MIR 5, de 4525±20 años AP, en la costa de Mar Chiquita (García et al., 2012; Fabra, 2014). Este linaje también se ha detectado en individuos de la región chaco-santiagueña[9], al norte, por lo cual se han propuesto relaciones biológicas entre poblaciones de ambas regiones (Tavella et al., 2020).
A partir de 2500 años AP se destacan las ocupaciones en los sitios El Diquecito (ED), la actual isla El Mistolar –sitios Rolo de Depetri (RD) y Mistolar (EMist) – y Estancia La Elisa (EE09). Hasta el momento, El Diquecito representa las primeras evidencias del asentamiento de poblaciones estables en el área, que utilizan de forma permanente y recurrentemente el espacio, y se vinculan con otras poblaciones también de entornos fluviales, del norte de la región pampeana (Prates & Bonomo, 2017). Las especies vegetales consumidas son Geoffroea decorticans, del género Prosopis y Trithrinax campestris(Tavarone et al., 2019). Es en este momento donde aparecen estructuras circulares de tierra cocida, las cuales tendrán continuidad en el registro arqueológico de los sitios hasta alrededor de 600 años AP (sitio Punta del Silencio –932/18PS–). Estas estructuras han sido consideradas “marcadores” de la presencia de grupos con economías cazadora-recolectora-pescadora en las llanuras de la región chaqueña, en el centro de Argentina, particularmente ríos y lagunas, desde la cuenca del Paraná hasta el piedemonte de las Sierras Pampeanas, y desde el norte de Buenos Aires hasta Mar Chiquita (Ceruti, 1998, 2000). En cuanto a la cerámica, si bien Bonofiglio (2010, 2011, 2015) la asocia a ocupaciones tardías, nuestras investigaciones en curso en el sitio El Diquecito podrían generar un cumulo de información para discutir la cronología asociada a estos desarrollos tecnológicos, al menos, hasta 2000 años AP.
Respecto a las afinidades biológicas, Tavella et al. (2020) destacan las similitudes para dos individuos –uno de la región chaco-santiagueña, al norte, y otro del sitio El Diquecito– que comparten el haplotipo C1d1b+16259 16271 16311, el cual no es compartido por otras 496 muestras antiguas, lo cual sugiere estrechos vínculos entre estas poblaciones. Para este linaje, considerando la antigüedad de la muestra del individuo S de dicho sitio (2331±46 14C años AP), se propone un origen local en el centro de Argentina. Este linaje se encuentra en la actualidad en mayor frecuencia en poblaciones del Gran Chaco. Por otro lado, el haplotipo C4c que presenta el individuo del sitio Estancia La Elisa (1890±49 años 14C años AP) es hasta el momento, el único en Sudamérica con ese haplotipo (Nores et al., 2016). Dicho individuo presenta mutaciones privadas idénticas a una muestra del Delta Superior del Paraná. Estos datos, sumados a la presencia de un individuo con haplotipo D1j en las muestras del Delta del Paraná (Nores et al., 2019) sugiere estrechos vínculos, al menos biológicos, entre poblaciones adscriptas a la entidad arqueológica Goya-Malabrigo y otras asentadas en la costa de Mar Chiquita. Estas afinidades habían sido sugeridas por estudios de rasgos epigenéticos y de variación craneofacial (Fabra, 2008;Fabra & Demarchi, 2013).
Las condiciones frías y secas de inicios del Holoceno tardío, con niveles bajos de la laguna, se alternaron con fases húmedas y cálidas, hasta que entre 1500-1000 años AP se producen dos momentos con balance hidrológico positivo, similares a las condiciones que presenta la laguna en la actualidad (Piovano et al., 2009). Es sugerente asociar estos cambios hacia condiciones más húmedas y cálidas con un incremento en la ocupación del territorio, un cambio en las estrategias de subsistencia y hasta en la composición biológica de las poblaciones, ocurridos alrededor de 1200 años AP como ha sido reseñado en este trabajo a partir de distintas líneas de evidencia. Los contextos arqueológicos de los sitios El Diquecito, Colonia Müller, Laguna del Plata, Marull, Orihuela e Isla Tigre darían cuenta de estos procesos. Para las Sierras Centrales, se ha planteado que el proceso de adopción de la práctica hortícola significó un cambio de importancia en distintos aspectos organizativos de estas sociedades, incluyendo un mayor sedentarismo, la incorporación de nuevas tecnológicas, y una diversificación e intensificación de elementos de la vida material, social e ideológica (Laguens & Bonnin, 2009). Este proceso de intensificación también se ha planteado para grupos con economía cazadora-recolectora en la región pampeana hacia momentos finales del Holoceno tardío, coincidente con la aparición en el registro arqueológico de la cerámica, y una mayor especificidad en la producción artefactual cerámica y lítica, diversificación en los recursos faunísticos aprovechados, un aumento en la densidad de población, variabilidad intra e intersitio, conjuntos artefactuales más formales y mayor equipamiento de sitios, solapamiento de espacios sagrados y domésticos, aparición de áreas formales de entierro, redundancia ocupacional, entre otras características (Martínez, 2017a).
Este proceso de intensificación social y productivo, que habría iniciado a partir de 1200 años AP con un momento de mayor expresión entre 700 y 500 años AP (sitios El Diquecito –ED08 y ED010–, El Mistolar –Emist–, Punta del Silencio –932/18PS–, Loma Alta –LAlta–, Orihuela –Orih–, Laguna de la Sal –LagSal–) , se habría caracterizado en la costa sur de Mar Chiquita por una ocupación más intensiva y redundante de los sitios, la incorporación complementaria a la caza, pesca y recolección del cultivo de porotos, zapallo y maíz, la producción cerámica, así como un paulatino aumento demográfico y cambio en la composición biológica de la población que podría estar relacionado con nuevos vínculos sociales. La construcción de las estructuras circulares de barro cocido (sea para almacenamiento o cocción de alimentos, las dos hipótesis planteadas hasta el momento) merece una consideración especial: si bien es sugerente pensarla en el marco de este proceso de intensificación, los sitios donde han sido relevados poseen fechados desde 2500 años AP a 500 años AP, en algunos casos se han detectado junto a entierros (individuo A, El Diquecito, 698±42 14C años AP; individuo 2, sitio Punta del Silencio, 690±50 14C años AP) o fueron rotos parcialmente al excavar la fosa para la inhumación (individuo L, El Diquecito, 937±150 14C años AP), lo cual hace pensar que no se encontraban en uso al momento de la apertura de la tumba. Es necesario profundizar el estudio de estas estructuras, tanto para definir la cronología de su uso como las funciones que pueden haber cumplido en este contexto regional particular.
Para la tecnología cerámica se ha planteado la producción y el estilo local de los distintos artefactos. Si bien Bonofiglio (2009) sugiere diferencias en los modos de hacer respecto a las modalidades identificadas en las comunidades serranas y otras de zonas llanas, como aquellas que ocuparon la cuenca media del rio Xanaes, no avanza en un planteo que permita ahondar en esas diferencias. Propone que las similitudes con otras comunidades del litoral se reducen a ciertos aspectos (técnica de manufactura de tipo espiralado, antiplásticos, acabados alisados y pulidos, formas predominantes de contorno simple, no restringido), particularmente la decoración de tipo incisión en surco rítmico. Esta técnica fue detectada en sitios del Paraná Medio e Inferior (Di Prado, 2018), así como en la cuenca inferior del rio Uruguay (Ottalagano & Castro, 2011; Ottalagano et al., 2015) y en la región pampeana, en el área de la Depresión del Salado (González & Frere, 2010). Otros objetos de hallazgo frecuente en esta área son las vasijas de pequeño tamaño o miniaturas (Figura 5F), similares a las halladas en contextos del Paraná medio y superior han sido asociadas a un uso ritual, en algunos casos vinculados al contexto mortuorio (Politis & Bonomo, 2018; Ottalagano, 2020). Es posible pensar entonces en un modo de hacer compartido con otras sociedades de las tierras bajas americanas, particularmente aquellas que vivieron en entornos fluviales o lacustres (Bonofiglio, 2011; Ceruti, 2000). Asociada a esta innovación, entre 1200 y 1000 años AP se habría establecido la práctica del cultivo a pequeña escala, particularmente de porotos, zapallo y maíz en el sitio El Diquecito (Tavarone et al., 2019). Tanto las cronologías como el complejo de plantas consumidas son parcialmente similares[10] y contemporáneas con los desarrollos documentados en el Delta del río Paraná (Bonomo et al., 2011b, 2017). Más allá de estas tradiciones compartidas a escala macroregional, coincidimos con Politis y colaboradores (2001) en que la adopción de cualquier tecnología se produce en contextos cultural e históricamente situados, mediados por relaciones sociales, y no pueden vincularse exclusivamente a cambios en las estrategias de subsistencia. En la región pampeana, la incorporación de la tecnología cerámica habría venido de la mano de otros cambios asociados a un incremento poblacional, cambios en los comportamientos mortuorios, un aumento en la producción y sus costos, un incremento en el componente vegetal de la dieta, aumento en la incorporación de carbohidratos a la dieta, diversificación y mayor consumo de taxa de tamaño pequeño, un incremento en los instrumentos de molienda y una reducción de la movilidad residencial (Politis et al., 2001; Barrientos & Perez, 2004; González, 2005; Martínez, 2008-2009; Gomez Otero & Novellino, 2011; Flensborg, 2013; Flensborg et al., 2013; Stoessel & Martínez, 2014; Martínez, 2017b, entre otros). En nuestra área, dicho proceso habría comenzado hacia 2000 años AP, y hacia 1200-1000 años AP estaría establecido, con un incremento paulatino en la señal arqueológica de los sitios. La presencia de grandes cantidades de tiestos cerámicos permiten pensar en ocupaciones permanentes[11] (Fabra et al., 2013). Si bien reconocemos las limitaciones que presentan los sitios de superficie como El Diquecito para establecer cronologías a las asociaciones contextuales, es interesante pensar en los procesos sociales de los cuales derivan prácticas como las alfareras, y vincularlos con estilos decorativos similares y tradiciones que se remontan a 1700-1500 años AP en sociedades cazadoras-recolectoras de la región pampeana (Politis et al., 2001).
Estos procesos de intensificación productiva y social también se reflejan en un uso más intensivo del cuerpo: las mayores prevalencias de cambios entesiales en los miembros inferiores a partir de 1200 años AP coinciden en cronología con la incorporación de la cerámica y las prácticas hortícolas, las cuales seguramente implicaron nuevas demandas y una diversificación en la cantidad o tipo de actividades realizadas. Hasta el momento, no hay indicios para pensar que este proceso de intensificación social se asocie a una creciente complejidad social: no hay cambios en las prácticas mortuorias, o en otros aspectos materiales de la cultura[12]. Las diferencias son tan notables en cuanto a las modalidades de inhumación entre las regiones serranas y llanas, particularmente aquellas vinculadas a ambientes lacustres, que podrían estar sugiriendo no sólo distintas concepciones en torno a la muerte sino también otras cosmovisiones y concepciones en torno a aspectos sociales, relacionales y simbólicos. Similitudes con sociedades vinculadas a entornos fluviales o llanos quedan evidenciadas en las prácticas mortuorias: a 100 km al sur de la laguna Mar Chiquita, en la cuenca del río Tercero que desemboca en el Paraná, se han reportado sitios monticulares, con inhumaciones de varios individuos, algunos de ellos pintados con ocre (Nimo, 1946), que asemejan a conjuntos recuperados en el delta del Paraná (Gaspary, 1950; Bonomo et al., 2011a; Scabuzzo et al., 2015; Kozameh et al., 2018, Castro & Castiñeira Latorre, 2018; Ramos van Raap, 2018) y región pampeana (Martínez et al., 2007). También la presencia de moluscos asociados a entierros puede sugerir tradiciones compartidas con poblaciones del noreste argentino (Lamenza et al., 2016; Ramos van Raap, 2018).
Con posterioridad al balance hidrológico positivo fechado en 1100 años AP, se produce un nuevo reverso hidrológico que continua con fases frías y secas hasta mediados del siglo XVIII. Interesa destacar que hace 834 años AP se daría el momento más extremo, con un descenso en el nivel de la laguna hasta 60,5 msnm, similar al observado hace 4700 años AP y hace 2000 años AP (Piovano com. pers.). Para esos momentos y hasta 500 años AP, las costas de la laguna alcanzarían la mayor ocupación del espacio por parte de poblaciones humanas, suponiendo mayor concentración poblacional en virtud de la intensificación de la señal arqueológica en ese período (Figura 4). Procesos similares de intensificación se han detectado en los ríos Paraná y Uruguay entre 800 y 500 años AP (Capdepont Caffa, 2018). El desmejoramiento de las condiciones climáticas habría generado una disminución en la disponibilidad de recursos, lo que sumado a una mayor densidad poblacional en el área explicaría el surgimiento de tensiones sociales y enfrentamientos intra o inter comunitarios, como el caso representado por los individuos 1 y 2 de la cuadrícula B del sitio El Diquecito (750±85 14C años AP), con lesiones óseas y un contexto mortuorio que ha sido vinculado a situaciones de violencia interpersonal (Fabra et al., 2015).
A partir de 500 años AP la señal arqueológica disminuye notablemente, con evidencias únicamente en los sitios Isla Orihuela y Río Dulce. La calibración de los fechados ubica a estas ocupaciones en los siglos previos a la conquista española; resta indagar si se trata de un sesgo de muestreo, o efectivamente se produjo una reducción o desplazamiento de la población.
En este trabajo se presentó una síntesis de las investigaciones bioantropológicas y arqueológicas llevadas adelante en la costa sur de la laguna Mar Chiquita. Esta área, particularmente las costas Este y Oeste de la Laguna del Plata, ha sido habitada por poblaciones humanas desde al menos 5000 años antes del presente (Fabra et al., 2014). Las investigaciones han puesto en evidencia una ocupación larga y relativamente estable –con un hiato entre 3800-2500 años AP–, caracterizada por la continuidad en los modos de vida, en estrecha vinculación con el ambiente lacustre que generan los humedales de la Laguna Mar Chiquita (Fabra, 2014). El aprovechamiento de las llanuras adyacentes a los ríos Suquía y Xanaes para la pesca o caza de especies de hábitos acuáticos ya había sido propuesta por Bonofiglio (2004, 2009, 2011, 2015), y más recientemente, estudios de isótopos estables han permitido sugerir el consumo de dietas mixtas (Laguens et al., 2009; Fabra & González, 2019), compuestas por vegetales silvestres y cultivados así como animales vinculados a ambientes lacustres. La arqueobotánica da cuenta de la continuidad en el uso de Geoffroea decorticans y del género Prosopis desde al menos 2500 años AP. La incorporación de vegetales cultivados tales como Phaseolus sp. alrededor de 1200 años AP, y posteriormente Cucurbita sp. y Zea mays hacia el 1000 AP fue complementaria a estrategias de caza, pesca y recolección de frutos silvestres (Tavarone et al., 2019).
Algunos elementos permiten distinguir a estas comunidades de las asentadas en regiones aledañas, particularmente serranas de la provincia: la forma de inhumar a los muertos es la que plantea mayores diferencias, siendo excluyente en las llanuras los entierros primarios extendidos, decúbito dorsal o ventral (en menor medida). Esta modalidad no ha sido registrada en las Sierras Centrales, pero si en las llanuras cercanas al rio Paraná (Ceruti, 2000). Si hay que mencionar no se han hallado hasta el momento evidencias de paquetes secundarios, uso de pigmentos o de termoalteración que son propios del Delta del Paraná (Ramos van Raap, 2018). También en el área de estudio, la presencia constante de ”hornitos” en el paisaje lacustre, así como elementos decorativos en vasijas, estatuillas (Figura 5G, J), o miniaturas (Figura 5F), la utilización de cestas y/o redes como soportes para la elaboración de contenedores cerámicos (Figura 5K), o la representación de ciertos animales (pájaros, particularmente), remiten a sociedades del litoral, particularmente el modelado de aves en Goya-Malabrigo y la región chaqueña (Figura 5D, E, H). Algunos artefactos, definidos como piedras con hoyuelos o placas grabadas en cerámica (Figura 5F), han sido asociados siguiendo a Serrano (1945) y Rodríguez (1992, citado por Bonofiglio, 2009) con la tradición Humaita de las tierras bajas sudamericanas (sur de Brasil, este de Paraguay, noreste de Argentina y norte de Uruguay).
En resumen, las investigaciones desarrolladas hasta el momento han permitido reafirmar vínculos biológicos y sociales con otras poblaciones, además de las asentadas en las Sierras Centrales, y comenzar a pensar en una dinámica poblacional de amplio rango, principalmente con otras comunidades de ambientes fluviales y lagunares. Se espera que la síntesis presentada en este trabajo permita contribuir al estudio de las sociedades humanas vinculadas a estos entornos, particularmente aquellos caracterizados por importantes y regulares fluctuaciones hidroclimáticas. Estas comunidades compartirían los criterios explicitados por Prates y Bonomo (2017) para sociedades relacionadas con ambientes acuáticos. La presencia de impresiones de redes y cestas en fragmentos cerámicos, así como los datos isotópicos permiten inferir el aprovechamiento de aves y peces, sin embargo, la intensidad del uso de dicho entorno no ha sido abordada. Resta avanzar sobre otros interrogantes, tales como la organización de las tecnologías cerámica y lítica, el uso del espacio y la movilidad de los grupos humanos, la importancia del consumo de especies de ambientes acuáticos a lo largo del tiempo, vínculos entre humanos y otros animales en aspectos relacionados tanto a su consumo como a esferas simbólicas, y el impacto en la salud de los cambios en las estrategias de subsistencia, entre otros.
A los Museos públicos de la región, que forman parte de la Asociación Civil Amigos del Patrimonio Histórico de Ansenuza, Suquía y Xanaes (APHA), por su enorme compromiso con el estudio, la difusión y la puesta en valor de las investigaciones sobre los pueblos originarios de la región: Carlos Ferreyra, Jose Luis Giraudo, Eduardo Villareal, Néstor Cocco, Luis María Cerda, Ulises Carrara, Angélica Montoya, del Museo Histórico Municipal de La Para. A Juan José Scienza, Hugo Giraudo del Museo de Ciencias Naturales de la región de Ansenuza “Anibal Montes” y a quienes nos acompañaron en gestiones anteriores de la institución: Alberto Rumachella, Laura Haas, Mercedes Sudar e Hilda Schwartz. A los integrantes del equipo, que hicieron posible los trabajos de campo y posteriores investigaciones, en vínculo con las comunidades locales: Mariela Zabala, Soledad Salega, Claudina González; Aldana Tavarone, Darío Ramírez, Luciano Loupias, Paloma Zarate, Romina Canova, Julieta Bellis, Lucia Andrade, Ana Paula Alderete y Eva Ferreyra. A Marcos Salvatore, por el mapa base confeccionado para la Figura 2. A Micaela Spangaro por las fotografías que forman parte de la figura 5. A Gisela Sario por los comentarios respecto al material lítico de la figura 5. A Eduardo Berberian, quien gentilmente autorizó la publicación del fechado realizado en el sitio Loma Alta. A Gustavo Politis y Guillermo Lamenza, por sus lecturas críticas y valiosos aportes al manuscrito, los cuales, junto a los realizados por un revisor, contribuyeron a mejorar sustancialmente el trabajo. Finalmente, a Marta Bonofiglio, un reconocimiento especial, por sus trabajos pioneros en la región, por haber sostenido que en las llanuras de la provincia de Córdoba “había indios”, y que sus sociedades merecían ser estudiadas en profundidad.
[1] Se tiene en cuenta la periodificación propuesta por Walker et al. (2019): Holoceno Medio -~8325-4250 años AP- y Holoceno Tardío -<4250 años AP.
[2] En ese trabajo se pudo estimar que los huesos wormianos sagital y epiptérico, así como la foramina cigomático facial tienen mayor peso en el PCI, y que el hueso wormiano epiptérico, la escotadura supraorbital y la foramina infraorbital múltiple son los rasgos no métricos con mayor peso en el PCII (Fabra et al., 2005).
[3] Los autores sugieren que la disminución en la talla observada en individuos femeninos, y el leve incremento en individuos masculinos no pueden ser explicados atendiendo a un único factor, más bien a una conjunción de causas entre las que pueden mencionarse además de las genéticas, el empeoramiento de las condiciones climáticas durante la Pequeña Edad de Hielo, la disminución en la disponibilidad de recursos, desmejoramiento de la salud (Loupias & Fabra, 2019).
[4] El fechado fue realizado sobre restos óseos del individuo recuperado en dicha cuadricula.
[5] Vale aclarar que salvo los casos mencionados en este trabajo, no se han hallado hasta el momento materiales asociados a los entierros humanos, sea como ajuar o acompañamiento funerario, en esta área.
[6] Vale aclarar que en dicho trabajo, las autoras agruparon las muestras de la costa sur de la Laguna Mar Chiquita junto con otras del Noreste, Sierras chicas y la Llanura oriental, todas subregiones de la provincia de Córdoba.
[7] La eburnación es un cambio asociado a la enfermedad degenerativa articular, definida por la apariencia suave y pulida de la superficie articular, y en grados severos, surcos (Rogers & Waldron, 1995).
[8] Hasta el momento, no se cuenta con restos arqueofaunísticos asociados a los sitios de este bloque temporal. Sí con datos arqueobotánicos, presentados y discutidos por Tavarone et al. (2019).
[9] Tavella et al. (2020) mencionan que la cronología para 8 de las muestras de los Bañados de Añatuya podría corresponder con la fase Oloma-Bajada-Icaño, de ca. 1400-1600 dC. Para las otras dos muestras, no tienen datos de procedencia o cronológicos precisos.
[10] En la costa sur de Mar Chiquita no se han encontrado evidencias hasta el momento de macro o microrestos de las familias Solanaceae, Chenopodiaceae, Polygonaceae, Poaceae (Oryzoideae) y Amaranthaceae (Sánchez et al., 2013; Colobig et al., 2015).
[11] Se hace referencia a los sitios Loma Alta (650±60 14C años AP, cantidad aproximada de fragmentos N= 850) –Bonofiglio, 2010–, Isla Orihuela costa sudeste (487±45 14C años AP, cantidad aproximada de fragmentos N= 250) –Fabra, 2011– y El Diquecito (537±57 14C años AP, cantidad aproximada de fragmentos N= 900) –Fabra et al., 2013–.
[12] Otro cambio que se puede mencionar entre momentos iniciales y finales del Holoceno Tardío es el de la práctica deformatoria del cráneo para algunos individuos, que pasó de ser de tipo circular a tabular erecta. La cronología de estas modalidades y sus implicancias en cuanto identidad individual o grupal, roles sociales o procesos vinculados con migraciones o contactos entre poblaciones será abordada en otro trabajo.
Figura 1.
Ubicación de los sitios arqueológicos analizados en este trabajo: 1. Miramar, 2. Estancia La Elisa, 3/19/20. La Para (aprox.), 4. Costa Este Laguna del Plata (aprox), 5. Costa oeste Laguna del Plata (aprox), 6. El Diquecito, 7. El Mistolar/Rolo de Depetri, 8. Colonia Muller, 9. Laguna del Plata (aprox), 10. Marull (aprox.), 11. Orihuela, 12. Isla Tigre (aprox), 13. Isla Orihuela, 14. Punta del Silencio, 15. Loma Alta, 17. Playa Grande, 18. Constantino, 21. Campo Bocassi-Agua Mansa, 22. Campo Milesi, 23. Tomasini-La Cerca, 24. Isla Larga, 25. La Loma/Campo Mare
Figura 2.
A. Sitio El Diquecito, Enero 2008; B. conjuntos de material cerámico en superficie, sitio El Diquecito, Abril 2008; C. Excavación entierro cuadricula Q, sitio El Diquecito Abril 2010; D. Excavación, sitio Isla Orihuela, Abril 2011; E. Excavación entierro 4, sitio Punta del Silencio, Agosto 2018; F. Prospección, estructura circular, sitio Rolo de Depetri, Septiembre 2018
Sitios arqueológicos prospectados y/o excavados en la costa sur de la Laguna Mar Chiquita, cantidad de restos humanos y materialidades asociadas. No incluye colecciones de museos regionales
Fechados radiocarbónicos obtenidos en la costa sur de la Laguna Mar Chiquita
Refs.: Codigo de Laboratorio: MTC (Escuela de Ciencias de Frontera, Universidad de Tokio, Japon), UCI (Universidad de Irving, Estados Unidos), AA (Universidad de Arizona, Estados Unidos), (Laboratorio de Tritio y Radiocarbono, Universdidad Nacional de La Plata, Argentina); Tipo: H: hueso humano, D: diente humano, M: malacologico; C: carbon. * Las calibraciones fueron realizadas con el programa OXCAL V4 3.2 (Ramsey 2017), IntCal 13 curva atmosferica (Reimer et al., 2013)
Figura 3.
Fechados radiocarbónicos calibrados para la costa sur de laguna Mar Chiquita
Figura 4.
Cantidad de fechados radiocarbónicos –sin calibrar–, considerando agrupaciones de 200 años
Figura 5.
A. Piedra grabada, posible afilador, sitio Loma Alta; B. punta de proyectil triangular apedunculada, sitio Loma Alta; C. raedera, sitio Loma Alta; D. aplique zoomorfo, sitio Loma Alta; E. aplique, representación de búho, sitio Loma Alta; F. Miniatura de vasija, sitio Loma Alta; G. borde con motivos incisos, de tipo surco rítmico punteado, sitio Loma Alta; H. vaso hueco zoomorfo, representación de ave, sitio Loma Alta; I. vasija entera, sitio Loma Alta; J. cuenco reconstruido, con incisiones y borde en ondas, sitio Loma Alta; K. fragmento con impresión de redes y cestas, pared interna, sitio El Diquecito; L. vasija con asa, Laguna del Plata; M. estatuilla antropomorfa, de tipo placa incisa, sitio Loma Alta.


