Resumen: Panochthus Burmeister constituye uno de los xenartros más abundantes, diversificados y ampliamente distribuidos del Pleistoceno sudamericano, como así también uno de los gliptodóntidos de mayor distribución latitudinal y altitudinal. En esta contribución se analizan aquellos registros asignables al género, efectuados en áreas de altura, en orden decreciente de altitud: (1) MURB 1906: ̴ 4000 m.s.n.m, Potosí (Bolivia); (2) MUSM 3632: ̴ 3800 m.s.n.m, Desaguadero (Perú); (3) MHNC-13491: ̴ 2600 m.s.n.m, Cochabamba (Bolivia); (4) MNPA-V 006598: ̴ 1870 m.s.n.m, Valle de Tarija (Bolivia); (5) MUFYCA 383: ̴ 920 m.s.n.m, Valle de Traslasierra (Argentina). Se discute la probable relación existente entre la altitud geográfica y la masa corporal de los ejemplares de P. intermedius Lydekker. En esta especie se observa que los individuos que habitan áreas de altura muestran valores de masa corporal significativamente menores que los de aquellos de la llanura pampeana. Esta relación también se registra en otros xenartros, así como en otros grupos de mamíferos. Estos taxones expresarían un patrón inverso de la regla de Bergmann. De esta manera, la “reducción de la disponibilidad de recursos” podría ser la razón ecológica que explicaría el menor tamaño de los individuos estudiados. Finalmente, P. hipsilis Zurita, Zamorano, Scillato-Yané, Fidel, Iriondo & Gillette es una especie endémica del Altiplano Boliviano y sus restos son los hallazgos de Panochthus y, posiblemente, de Glyptodontidae realizados a mayores alturas.
Palabras clave: Mamíferos, Panochthus, Altitud, Pleistoceno, América del Sur.
Abstract: Panochthus Burmeister is one of the most abundant, diversified and widely distributed xenarthrans of the South American Pleistocene, as well as one of the glyptodontids with higher latitudinal and altitudinal distributions. This contribution analyzes those records assigned to the genus, in decreasing order of elevation: (1) MURB 1906: ̴ 4000 masl, Potosí (Bolivia). (2) MUSM 3632: ̴ 3800 masl, Desaguadero (Perú). (3) MHNC-13491: ̴ 2600 masl, Cochabamba (Bolivia). (4) MNPA-V 006598: ̴ 1870 masl, Tarija Valley (Bolivia). (5) MUFYCA 383: ̴ 920 masl, Traslasierra Valley (Argentina). We discuss the probable relationship between geographic altitude and body mass of P. intermedius Lydekker specimens. In this species, the individuals that inhabited high areas show significantly lower values of body mass than those fromthe Pampas plains. This relationship has also been recorded in other xenarthrans, as well as in other groups of mammals. These taxa could express an inverse Bergmann's rule pattern. In this sense, a “reduction in the availability of resources” could be the ecological reason to account for the smaller size of the individuals studied. P. hipsilis Zurita, Zamorano, Scillato-Yané, Fidel, Iriondo & Gillette is an endemic species fromthe Bolivian Altiplano; its findings represent thegreatest altitudinal records for Panochthus and possibly for the Glyptodontidae.
Keywords: Mammals, Panochthus, Altitude, Pleistocene, South America.
Resumo: Panochthus Burmeister é um dos xenartros mais abundantes, diversificados e amplamente distribuídos do Pleistoceno sulamericano, bem como um dos gliptodontídeos de maior distribuição latitudinal e altitudinal. Nesta contribuição analisam-se os registros atribuíveis ao gênero, feitos em áreas de elevada altitude, em ordem decrescente: (1) MURB 1906: 4000 msnm, Potosí (Bolívia); (2) MUSM 3632: 3.800 msnm, Desaguadero (Peru); (3) MHNC-13491: 2.600 msnm, Cochabamba (Bolívia); (4) MNPA-V 006598: 1.870 msnm, Valle de Tarija (Bolívia); (5) MUFYCA 383: 920 msnm, vale de Traslasierra (Argentina). Discute-se a provável relação entre a altitude geográfica e a massa corporal dos espécimes de P. intermedius Lydekker. Nesta espécie observa-se que os indivíduos que habitam áreas de elevada altitude apresentam valores de massa corporal significativamente menores que aqueles da planície pampeana. Esta relação também é registrada em outros xenartros, bem como em outros grupos de mamíferos. Estes táxons expressariam um padrão inverso à regra de Bergmann. Desta maneira, a “redução da disponibilidade de recursos” poderia ser a razão ecológica que explicaria o menor tamanho dos indivíduos estudados. Finalmente, P. hipsilis Zurita, Zamorano, Scillato-Yané, Fidel, Iriondo & Gillette é uma espécie endêmica do Altiplano boliviano e seus restos são os achados de Panochthus e, possivelmente, de Glyptodontidae provenientes de altitudes mais elevadas.
Palavras-chave: Mamíferos, Panochthus, Altitude, Pleistoceno, América do Sul.
Artículo
Restos de †Panochthus Burmeister (Xenarthra; Cingulata; Glyptodontidae) registrados en áreas de altura: descripción y comentarios
Remains of †Panochthus Burmeister (Xenarthra; Cingulata; Glyptodontidae) recorded at high altitudes: description and comments
Restos de †Panochthus Burmeister (Xenarthra; Cingulata; Glyptodontidae) registrados em áreas de altitude: descrição e comentários
Recepción: 01 Enero 2020
Aprobación: 03 Julio 2020
Publicación: 20 Julio 2020
Los xenartros constituyen un grupo muy particular de mamíferos placentarios y característico de la Región Neotropical. Se encuentran ampliamente representados en el registro fósil sudamericano, tanto por su extensión temporal como por su frecuencia (Paula Couto, 1979; Scillato-Yané 1982; Wetzel, 1982; Zamorano, 2012, 2013; Gaudin & Croft, 2015). La mayoría de sus taxones provienen de América del Sur (Paleoceno tardío-Actualidad) y, en menor medida, de América del Norte y América Central (Mioceno tardío-Actualidad) (Carlini & Zurita, 2010). La diversificación del grupo se ve reflejada en la amplia gama de dietas, desde omnívoros generalizados (con una amplia diversidad de hábitos alimenticios, carroñeros, predadores, hasta la elevada especialización animalívora de los Myrmecophagidae) a herbívoros ramoneadores o pastadores, de hábitos selectivos o indiscriminados (Bargo & Vizcaíno, 2008; Vizcaíno et al., 2006, 2008; Vizcaíno, 2009; Zamorano et al., 2018); la heterogeneidad de ambientes que ocupan (áreas abiertas, arboladas, áridas-semiáridas, y diferentes tipos de pastizales, desde fríos a cálidos y subtropicales [Scillato-Yané et al., 1995], como asimismo, en el gran rango de altitudes en las que se encuentran (Pujos, 2006, 2008; Zurita et al., 2012; Zurita et al., 2017).
Los Cingulata (armadillos y gliptodontes) son los xenartros más antiguos en el registro y están representados por cinco grandes agrupaciones: Dasypodidae, Chlamyphoridae, Pampatheriidae, Pachyarmatheriidae y Glyptodontidae (ver Gibb et al., 2016; Fernicola et al., 2018). Glyptodontidae representa uno de los clados más emblemáticos y de mayor éxito evolutivo dentro de los cingulados fósiles (Zurita et al., 2016). En su extensa historia evolutiva (Eoceno tardío-Holoceno temprano), tanto el registro más antiguo (McKenna & Bell, 1997) como el más moderno (Gutiérrez et al., 2010) corresponden al actual territorio argentino, habiendo alcanzado una diversidad notable y tamaños a veces gigantescos (Scillato-Yané & Carlini, 1998; Fariña, 2001; Zamorano et al., 2013, 2014a), pudiendo superar los 2300 kg (Soibelzon et al., 2012).
Panochthus Burmeister es uno de los gliptodóntidos más abundantes, diversificados y ampliamente distribuido del Pleistoceno sudamericano, como así también uno de los cingulados de mayores dimensiones (Zamorano, 2012; Zamorano et al., 2014a, 2014b, 2015). Diversas contribuciones coinciden en que se trata de un género monofilético (Zamorano, 2012; Zamorano & Brandoni, 2013; Zurita et al., 2017). La última revisión del género (Zamorano et al., 2014a) reconocía seis especies, a las que recientemente se ha agregado una séptima (Zurita et al., 2017), estas incluyen: (1) P. intermedius Lydekker, Ensenadense (Pleistoceno temprano tardío) de la ciudad de Buenos Aires (Argentina) y Cochabamba (Boliva); (2) P. subintermedius Castellanos, Ensenadense (Pleistoceno temprano tardío) de la ciudad de Buenos Aires (Argentina); (3) P. tuberculatus (Owen), Bonaerense (Pleistoceno medio) y Lujanense (Pleistoceno tardío) de gran parte de Argentina, Uruguay, sur y centro de Bolivia y sur de Brasil; (4) P. frenzelianus Ameghino, Bonaerense (Pleistoceno medio) de la provincia de Buenos Aires (Argentina) y los alrededores de Montevideo (Uruguay); (5) P. greslebini Castellanos, Lujanense (Pleistoceno tardío) de la provincia de Buenos Aires (Argentina) y Pleistoceno (sensu lato) del noreste de Brasil; (6) P. jaguaribensis (Moreira), Pleistoceno (sensu lato) del noreste de Brasil; y (7) P. hipsilis Zurita, Zamorano, Scillato-Yané, Fidel, Iriondo & Gillette, Pleistoceno (sensu lato) de la Cordillera Oriental de Bolivia. Junto con Glyptodon Owen (Glyptodontinae, Glyptodontini), Panochtus es uno de los gliptodontes de mayor distribución latitudinal y altitudinal en América del Sur (Zamorano & Jara Almonte, 2018; Zurita et al., 2009, 2016).
En esta contribución se analizan aquellos materiales asignables a Panochthus registrados en áreas de altura, detallándose en cada caso su clasificación sistemática y procedencias geográfica y estratigráfica; asimismo, se realiza una breve descripción y comentarios de cada ejemplar. Por otra parte, se discute la posible relación existente entre las variaciones de la masa corporal y la altitud en los taxones tratados.
La mayoría de los materiales analizados fueron dados a conocer en trabajos previos [MHNS 8, MURB 1906A y MURB 1906B en Zurita et al. (2017); MHNC-13491 en Zurita et al. (2011); MNPA-V 006598 en Zurita et al. (2009); MUFyCA 383 en Castellano (1942); MUSM 3632 en Zamorano & Jara Almonte (2018)]; por otro lado, la publicación que describe minuciosamente a UATF s/n está siendo actualmente preparada. Para la sistemática se siguió el esquema propuesto recientemente por Zamorano (2019). Los ejemplares analizados fueron comparados solamente con P. tuberculatus (especie tipo), P. hipsilis y P. intermedius (las dos últimas son aquí estudiadas), con el fin de evitar extender innecesariamente esta contribución. Todos los valores incluidos en la Tabla 1 se expresan en milímetros (mm), con un margen de error de 1 mm (Fig. 1).
Medidas de las especies mencionadas
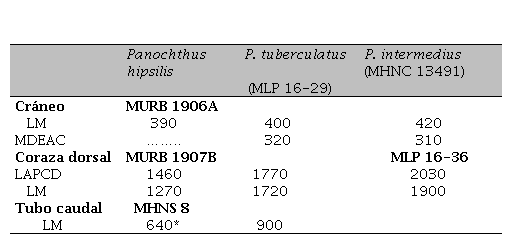
Abreviaturas: LAPCD= longitud antero-posterior siguiendo la curva dorsal; LM= longitud máxima en línea recta; MDEAC= máxima distancia entre arcos cigomáticos.
Referencias. *le falta el extremo distal.
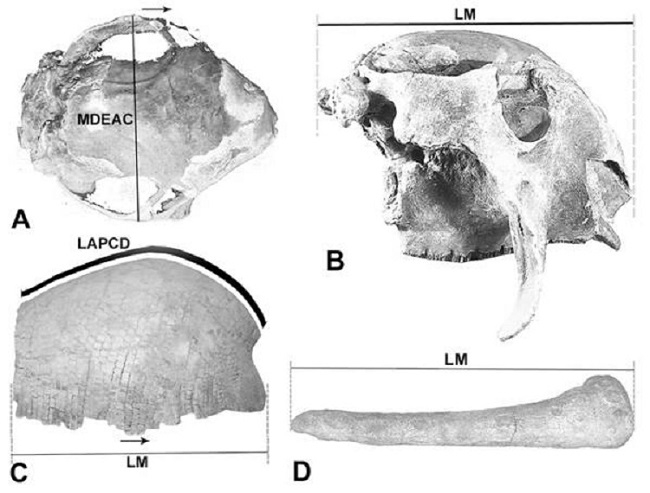
Figura 1.
Medidas calculadas.
A, vista dorsal del cráneo; B, vista lateral derecha del cráneo; C, vista lateral derecha de la coraza dorsal; D, vista lateral derecha del tubo caudal. Abreviaturas. LAPCD= longitud antero-posterior siguiendo la curva dorsal; LM= longitud máxima en línea recta; MDEAC= máxima distancia entre arcos cigomáticos.
MHNC: Museo de Historia Natural “Alcide d'Orbigny”; Cochabamba, Bolivia. MHNS: Museo de Historia Natural de Sucre; Sucre, Bolivia. MLP: División Paleontología Vertebrados, Facultad de Ciencias Naturales and Museo, Universidad Nacional de La Plata, Argentina. MNPA-V: Museo Nacional Paleontológico – Arqueológico; Tarija, Bolivia (colección vertebrados). MUFyCA: Museo Universitario Florentino y Carlos Ameghino, Rosario, Argentina. MURB: Museo Universitario “Ricardo Bohorquez”, Potosí, Bolivia. MUSM: Museo de Historia Natural de la Universidad Mayor de San Marcos, Lima, Perú. UATF: Universidad Autónoma “Tomás Frías,” Potosí, Bolivia.
Superorden XENARTHRA Cope, 1889 Orden CINGULATA Illiger, 1811 Suborden GLYPTODONTIA Ameghino, 1889 Superfamilia GLYPTODONTOIDEA Gray, 1869 Familia GLYPTODONTIDAE Gray, 1869 Género Panochthus Burmeister, 1866 Especie tipo. Glyptodon tuberculatus Owen, 1845 Panochthus hipsilis Zurita, Zamorano, Scillato-Yané, Fidel, Iriondo & Gillette, 2017. MURB 1906A: cráneo incompleto, y MURB 1906B: dos vértebras, coraza dorsal casi completa, con osteodermos asociados que pertenecen principalmente a la región lateral y otros restos indetermibados (Fig. 2B). Procedencia geográfica y estratigráfica: Pleistoceno tardío de inmediaciones inmediaciones de la ciudad de Potosí (Bolivia). Altitud aproximada: 4000 m.s.n.m (Fig. 3).
Breve descripción El cráneo es de morfología similar al de la especie tipo y al de P. intermedius, pero de menor tamaño. Como en P. tuberculatus y a diferencia de P. intermedius (holotipo) la coraza dorsal está elevada anteriormente, es de menor tamaño que la de P. tuberculatus y menor aún que aquella del holotipo de P. intermedius (ver Tabla1). Ornamentación de la coraza dorsal completamenmte reticular, excepto en la penúltima hilera de osteodermos adyacentes a la escotadura caudal y en los osteodermos mas latero-ventrales que muestran una figura central, de manera semejante a P. tuberculatus y a diferencia de P. intermedius,que retiene las regiones antero-dorsal y lateral, osteodermos con figura central. . MHNS 8: tubo caudal casi completo, al que le falta el extremo distal (Fig. 2A). Procedencia geográfica y estratigráfica: Pleistoceno tardío de las cercanias de la ciudad de Potosí (Bolivia). Altitud: 4000 m-s.n.m (Fig. 3).
Breve descripción Pese a estar incompleto por no preservarse el extremo distal, puede estimarse que el tubo caudal de P. hipsilis sería más corto que los de P.tuberculatus y P. intermedius (ver Tabla 1). Su ornamentación tiene mayoritariamente un patrón "en roseta", de manera semejante a P. intermediusy a diferencia de P. tuberculatus, exhibe un patrón reticulado en la mayoría de su superficie. Comentarios (MURB 1906A/1906B y MHNS 8) P. hipsilis, la especie más pequeña del género, es conocida solo por los materiales arriba descriptos. MURB 1906A/1906B es el holotipo de la especie, en tanto que MHNS 8 se atribuyó a esta especie principalmente porque posee la misma procedencia geográfica y estratigráfica, y porque al igual que en Propanochthus bullifer y en las demás especies Panochthus, existe una clara relación entre la ornamentación de la superficie de la coraza dorsal y aquella de la escultura externa del tubo caudal. Panochthus intermedius Lydekker, 1895 . MHNC-13941: ejemplar casi completo, del que se han preservado cráneo y mandíbula, parte del esqueleto postcraneal, escudete cefálico, coraza dorsal casi completa aunque sumamente fragmentada (falta la parte de la región postero-dorsal), fragmentos de algunos anillos caudales y porción proximal del tubo caudal (Fig.2F) (Zurita et al., 2011). Procedencia geográfica y estratigráfica: Formación Sacaba (Pleistoceno medio-tardío [ver Soruco & Díaz Martinez, 1996] de Cochabamba, Bolivia. Altitud: 2600 m.s.n.m (Fig. 3). Breve descripción El ejemplar aquí examinado es claramente de menor tamaño que el holotipio (MLP 16-36, coraza dorsal casi completa) (ver Tabla 1). Procedencia estratigráfica y geográfica del holotipo según Lydekker(1895, p.34). ·Pampeano del Puerto de Buenos Aires". Ensenadense (Pleistoceno temprano medio) del puerto de Buenos Aires (actualmente " Ciudad autónoma de Buenos Aires") Argentina. El cráneo es semejante en forma y tamaño a aquellos de P. tuberculatus y P. hipsilis. Osteodermosde la coraza dorsal con figura central distinguible, rodeada de figuritas periféricas en las regiones antero-dorsal, postero-dorsal y lateral, morfología que los diferencia de los P. tuberculatus y P. hipsilis, en las que su ornamentación es mayormente reticular. Comentarios La coraza dorsal (MHNC-13491) actualmente está fragmentada en asociaciones de osteodermos, pero fue descubierta casi completa. Comparada con el holotipo de P. intermedius (MLP-16-36), la morfología de la superficie expuesta de los osteodermos en las diferentes regiones es prácticamente idéntica, y difiere de todas las especies de Panochthus. El tamaño de ese ejemplar es sensiblemente menor al del holotipo, similar al de P. tuberculatus (ver Zamorano, 2014a) y mayor que P. hipsilis.

Figura 2.
Materiales de Panochthus analizados en la presente contribución. A, MHNS 8 (P. hipsilis) (modificado de Zurita et al., 2017); B, MURB 1906A/1906B (holotipo de P. hipsilis) (modificado de Zurita et al., 2017); C, MUSM 3632 (Panochthus sp.); D, MUFyCA 383 (Panochthus sp.); E, MNPA-V 006598 (Panochthus sp.); F, MHNC-13491 (P. intermedius) (modificado de Zurita et al., 2011).
Panochthus sp. . MNPA-V 006598: un osteodermo aislado de la coraza dorsal (del cual, lamentablemente, no existe registro fotográfico) y tres fragmentos del tubo caudal (Fig. 2E). Procedencia geográfica y estratigráfica: Pleistoceno (sensu lato) del valle de Tarija, Bolivia. Altitud: 1870 m.s.n.m. (Fig.3). Breve descripción El osteodermo corresponde a la región medio-dorsal; la superficie expuesta está cubierta completamente con figuritas tuberculares que siguen un claro patrón reticular. Dos fragmentos del tubo caudal corresponden a la región lateral, que muestra grandes figuras subelípticas rugosas; el fragmento restante, pertenece a la región ventral y posee figuritas tuberculares y figuras centrales. Comentarios Zurita et al. (2009) examinaron este espécimen y concluyeron que las características de MNPA-V 006598 permiten asignar a este como Panochthus sp., esto modificó la asignación original de Hoffstetter (1964:131-132), quien había atribuido este ejemplar a Panochthus cf. P. tuberculatus.

Figura 3.
Distribución geográfica de las especies de Panochthus y rango decreciente de altitudes de los registros obtenidos a mayores alturas que 900 m.s.n.m. A, Distribución geográfica; B, localidades donde fueron recuperados los materiales a mayor altitud; C, Histograma que muestra las altitudes de los registros de; a, alrededores de Potosí (holotipo de P. hipsilis) a ̴ 4000 m; b, Desaguadero, Perú (atribuido a Panochthus sp.) a ̴ 3800 m; c, cercanías de Cochabamba, Bolivia (atribuido a P. intermedius) a ̴ 2600 m; c, Valle de Tarija, Bolivia (atribuido a Panochthus sp.) a ̴ 1870 m; e, Valle de Traslasierra, Nono, Argentina (atribuido a Panochthus sp a ̴ 920 m (modificado de Zamorano & Jara Almonte, 2018).
. MUFyCA 383; tubo caudal completo, con restos del anillo proximal (Fig. 2D). Procedencia geográfica y estratigráfica:Pleistoceno (sensu lato) de las barrancas de Pizarro, margen derecho del río Grande de Nono,Valle de Traslasierra, provincia de Córdoba, Argentina. Altitud: 920 m.s.n.m (Fig. 3).
Breve descripción Este tubo es más largo que el de P. hipsilis, levemente más corto que el de P. tuberculatus y mucho más que el de P. intermedius (ver Tabla 1). Proximalmente es cónico-cilíndrico y deprimido en la porción distal. En el extremo distal tiene una sola figura apical (ver Zamorano et al., 2015), a diferencia de P. intermedius, P. hipsilis y de la mayoría de los materiales atribuidos a P. tuberculatus. La ornamentación predominante en el tubo caudal posee patrón “en roseta”, aunque en algunos sectores prevalece el patrón reticular.
Comentarios Este material fue asignado por Castellanos (1942, 1944) y Cruz (2011) a P. frenzelianus; sin embargo, Ameghino (1889) reconoció a Panochthus frenzelianus basándose en un cráneo, mandíbula, casquete cefálico, osteodermos de la región posterior del caparazón y anillos caudales (ver Fernicola et al., 2014). Zamorano (2012) decide no considerar al material MUFyCA 383 como parte del hipodigma de P. frenzelianus. Finalmente, Cruz (2013) lo designa como Panochthus sp.
· MUSM 3632: osteodermo aislado de la coraza dorsal (Fig. 2C). Procedencia geográfica y estratigráfica: Pleistoceno (sensu lato) de la ribera del río Desaguadero, departamento de Puno, provincia de Chucuito, Perú. Altitud: ~ 3800 m.s.n.m (Fig.3).
Breve descripción Este osteodermo presenta un contorno cuadrilátero trapezoide. Su grosor es menor a aquel de las especies de Glyptodon, y superior a las de Neosclerocalyptus Paula Couto (ver Zurita, 2007). La superficie dorsal consiste en tubérculos poligonales pequeños y similares entre sí en tamaño, distribuidos por toda la superficie, como en la mayoría de las regiones de la coraza dorsal de P. tuberculatus y P. hipsilis, y solo en los osteodermos de las regiones que no tienen figura central de P. intermedius (ver Zamorano et al., 2014a).
Comentarios Hoffstetter (1973) registra para Ymata (Puno) cf. Panochthus; aunque no brinda datos de este material. Pujos & Salas (2004) realizan una revisión de los registros de xenartros del Perú, mencionando el registro de Hoffstetter (1973) como cf. Panochthus, pero sin especificar tampoco el material. Asimismo, Scillato-Yané et al. (1995) indican que Perú forma parte de la distribución geográfica de Panochthus, sin justificar la mención. Scillato-Yané (com. pers.) manifestó que posiblemente dicho dato haya sido producto de una conversación con Hoffstetter. Este último también explica el motivo de la mención de Perú para la distribución de Panochthus en Zamorano et al. (2014a), trabajo en el cual Scillato-Yané es coautor. Finalmente, Zamorano & Jara Almonte (2018) confirman el registro y brindan detalles de la identificación taxonómica.
Discusión y conclusiones
Un aspecto interesante a discutir es la probable relación existente entre la altitud geográfica y la masa corporal de los ejemplares de P. intermedius analizados. En este sentido, se aprecia que MHNC-13491, proveniente de áreas de altura (Cochabamba), es significativamente menor al hallado en la llanura pampeana (Buenos Aires), MLP 16-36. El material proveniente de las áreas de altura posee una talla similar a la de la especie tipo, mientras que el holotipo es el espécimen de mayor tamaño documentado para el género (Zamorano et al., 2014a) (ver Tabla 1). Esta relación fue señalada también para otros xenartros. Pujos (2008) afirma que los Megatheriinae registrados en altitudes elevadas, excepto Megatherium (Pseudomegatherium) celendinense Pujos (una especie de Megatherium, particularmente grande, de los Andes del Norte de Perú [ver Pujos, 2006]), son de menor tamaño que los de altitudes bajas. En gliptodóntidos, Zurita et al. (2012) afirman que algunos ejemplares de Glyptodon restringidos a las áreas andinas son de menor tamaño que aquellos presentes en la región Pampeana. Asimismo, la reducción de la talla corporal con la altitud ha sido registrada también en otros linajes de mamíferos placentarios, como milodóntidos (Ameghino, 1899; McDonald et al., 2000), équidos (Alberdi et al., 1995; Alberdi & Prado, 2004) y gonfotéridos (Prado et al., 2005). Como se ha visto en otros grupos (e.g., Tárraga, 2009; Sargis et al., 2018) los taxones mencionados siguen un patrón ecogeográfico inverso a la clásica regla de Bergmann (Bergmann, 1847, pero ver Blackburn et al., 1999), regla para la que se han propuesto diversas modificaciones (e.g., Mayr, 1956; James, 1970; Gaston et al., 2008; McNab, 2010; Gutiérrez-Pinto et al., 2014). Dentro de los xenartros, McDonald et al. (2004) proponen que Paramylodon harlani (Owen) sigue la regla de Bergmann, en tanto que Brandoni et al. (2010) consideran que el menor tamaño de Mylodon darwini Owen, del Pleistoceno tardío de la Mesopotamia (Argentina), puede atribuirse a la regla de Bergmann. Sin embargo, el valor de dicha regla es discutido por muchos autores (e.g., Geist, 1987; Meiri & Dayan, 2003; Blackburn & Hawkins, 2004; Brehen & Fiedler, 2004; Meiri et al., 2007; Rodriguez et al., 2008; Barros et al., 2014; Brown et al., 2017). De acuerdo con las últimas publicaciones citadas, una de las principales críticas a la regla de Bergmann es que el incremento de volumen no significa directamente la eficaz regulación de la temperatura, sino que supone una mayor pérdida total de calor corporal, además de una mayor demanda energética (es necesaria una mayor ingesta de alimento). De este modo, la masa corporal podría descender en áreas de altura cuando el alimento se convierte en un factor limitante. De esta manera, la “reducción de la disponibilidad de recursos” (véase Geist, 1987; McNab, 2010) podría ser la razón ecológica que explicaría el menor tamaño de los individuos aquí estudiados comparados con otros estrechamente emparentados (cogenéricos o coespecíficos).
La evidencia paleontológica disponible sugiere que P. hipsilis es una especie endémica del Altiplano Boliviano (inmediaciones de Potosí), y que sus restos constituyen los hallazgos asignables a Panochthus y, posiblemente, a toda la familia Glyptodontidae, efectuados a mayores alturas (~4000 m.s.n.m). Esta especie debe sumarse a otras especies de la megafauna sudamericana que fueron propios de la región subandina: Cuvieronius hyodon (Ficher) (Proboscidea, Gomphotheriidae) (Prado et al., 2005); Hippidion saldiasi (Roth), Equus (A.) andium Branco, Equus (A.) insulatus (Ameghino), Equus (A.) santaeelenae (Spillman) y Equus (A.) lasallei (Perissodactyla, Equidae) (Alberdi & Prado, 2004).
A modo de síntesis, los registros fehacientes de materiales asignados a Panochthus se disponen en un orden decreciente de altura: (1) el holotipo de P. hipsilis (MURB 1906A/1906B) y un tubo caudal (MHNS 8), asignados como de P. hipsilis, procedentes de los alrededores de Potosí (Bolivia), a ̴ 4000 m.s.n.m; (2) un osteodermo aislado (MUSM-s/n) atribuido a Panochthus sp., de Desaguadero (Perú), a ̴ 3800 m.s.n.m; (3) el ejemplar casi completo (MHNC-13491) asignado a P. intermedius, de las cercanías de Cochabamba (Bolivia), a ̴ 2600 m.s.n.m; (4) un osteodermo y porciones del tubo caudal (MNPA-V 006598) atribuidos a Panochthus sp., del Valle de Tarija (Bolivia), a ̴ 1870 m.s.n.m. Finalmente, uno de los registros m.s.n.m hallados a mayor altitud en Argentina corresponde a un tubo caudal completo (MUFyCA 383) asignado a Panochthus sp., proveniente del Valle de Traslasierra, Nono, provincia de Córdoba, a ̴ 920 m.s.n.m.
A Alfredo E. Zurita por el envío de fotografías de material hallado en Bolivia. Al revisor (anónimo) y al editor (Dr. Edgardo Ortíz-Jaureguizar) por enriquecer notablemente esta publicación.
Medidas de las especies mencionadas
Abreviaturas: LAPCD= longitud antero-posterior siguiendo la curva dorsal; LM= longitud máxima en línea recta; MDEAC= máxima distancia entre arcos cigomáticos.
Referencias. *le falta el extremo distal.Figura 1.
Medidas calculadas.
A, vista dorsal del cráneo; B, vista lateral derecha del cráneo; C, vista lateral derecha de la coraza dorsal; D, vista lateral derecha del tubo caudal. Abreviaturas. LAPCD= longitud antero-posterior siguiendo la curva dorsal; LM= longitud máxima en línea recta; MDEAC= máxima distancia entre arcos cigomáticos.
Figura 2.
Materiales de Panochthus analizados en la presente contribución. A, MHNS 8 (P. hipsilis) (modificado de Zurita et al., 2017); B, MURB 1906A/1906B (holotipo de P. hipsilis) (modificado de Zurita et al., 2017); C, MUSM 3632 (Panochthus sp.); D, MUFyCA 383 (Panochthus sp.); E, MNPA-V 006598 (Panochthus sp.); F, MHNC-13491 (P. intermedius) (modificado de Zurita et al., 2011).
Figura 3.
Distribución geográfica de las especies de Panochthus y rango decreciente de altitudes de los registros obtenidos a mayores alturas que 900 m.s.n.m. A, Distribución geográfica; B, localidades donde fueron recuperados los materiales a mayor altitud; C, Histograma que muestra las altitudes de los registros de; a, alrededores de Potosí (holotipo de P. hipsilis) a ̴ 4000 m; b, Desaguadero, Perú (atribuido a Panochthus sp.) a ̴ 3800 m; c, cercanías de Cochabamba, Bolivia (atribuido a P. intermedius) a ̴ 2600 m; c, Valle de Tarija, Bolivia (atribuido a Panochthus sp.) a ̴ 1870 m; e, Valle de Traslasierra, Nono, Argentina (atribuido a Panochthus sp a ̴ 920 m (modificado de Zamorano & Jara Almonte, 2018).
. MUFyCA 383; tubo caudal completo, con restos del anillo proximal (Fig. 2D). Procedencia geográfica y estratigráfica:Pleistoceno (sensu lato) de las barrancas de Pizarro, margen derecho del río Grande de Nono,Valle de Traslasierra, provincia de Córdoba, Argentina. Altitud: 920 m.s.n.m (Fig. 3).


