1. Introdução
Nos últimos milhares de anos, a expansão das florestas em áreas de savanas e campos ocorreu em muitas partes da América do Sul, um fenômeno que tem sido geralmente atribuído às flutuações climáticas (Bueno et al. 2017; Arruda et al., 2018). No entanto, mudanças climáticas não são a única influência importante no limite destes ecótones. Outros processos, como a dispersão de espécies, características dos solos e distúrbios antrópicos também podem influenciar a distribuição da vegetação (Souza et al. 2020). Na Reserva da Biosfera da Serra do Espinhaço (RBSE), sudeste do Brasil, é possível verificar um complexo mosaico fitofisionômico decorrente de elevada heterogeneidade ambiental (i.e. variações climáticas, hidrológicas, edáficas e altitudinais) (Braga et al. 2017)
Na RBSE um sistema peculiar de floresta-campo merece destaque. Trata-se de “ilhas” de florestas estacionais naturais conhecidas como Matas de Galerias ou “Capões” (Coelho et al., 2018; Gonçalves et al., 2020). Constituem-se importantes centros de biodiversidade multitaxonômicos, apesar de ocuparem pequena extensão territorial (Fonseca et al., 2018), além de grande relevância na provisão de serviços ambientais e ecossistêmicos em múltiplas formas (Oliveira et al., 2015), especialmente no fornecimento de água (Mendonça-Galvão et al., 2011). Em todas as estações do ano, possuem, na maioria das amostragens, espécies típicas de áreas úmidas e alagadas (Silva et al., 2016).
Estes capões, comuns nas zonas de transição entre os biomas Mata Atlântica, Cerrado, Pantanal e Caatinga (Ribeiro e Walter, 2008), se enquadram como florestas estacionais do Domínio Atlântico e ocorrem associadas a uma transição brusca para áreas campestres da savana (Silveira et al., 2016), e seus limites são controlados pelo gradiente de fertilidade do solo e condições microclimáticas específicas (Coelho et al., 2018). Constituemformações florestais naturais que ocorrem em regiões deprimidas da paisagem e sob afloramento do lençol freático, que favorecem sua condição de alagamento mesmo em períodos de seca (Ribeiro e Walter, 2008; Fontes e Walter, 2011; Fontes et al., 2015).
A despeito de sua importância, estes ambientes constituem uma das formações mais ameaçadas da RBSE (Silva et al., 2016), seja pela forte atuação antrópica (Queiroz et al., 2018) e pela alta sensibilidade ao fogo (Alvarado et al., 2017), como pelo isolamento físico natural. Além disto, sua vegetação é composta por poucas espécies conspícuas, que necessitam de uma série de adaptações e atributos que permitam seu estabelecimento (Coelho et al., 2018). Assim, torna-se urgente a necessidade de preservar as mesmas, sendo apontadas como áreas prioritárias para conservação (Fonseca, 2020).
No entanto, ainda há muita negligência sobre esta fitocenose, sendo necessário pesquisas pontuais para enaltecer sua importância. Assim, o objetivo deste estudo foi realizar a caracterização florística de capões de mata (ilhas florestais) localizados em Unidades de Conservação da Reserva da Biosfera da Serra do Espinhaço. Esperamos que este artigo possa contribuir com o conhecimento da biodiversidade valiosa destes ambientes, servindo de subsídio para o estabelecimento de políticas públicas voltadas à sua conservação.
2. Material e métodos
2.1 Caracterização
da área de estudo
O estudo foi realizado no Parque Estadual do Biribiri (PEBi) e Parque Nacional das Sempre Vivas (PNSV), localizados na porção da Serra do Espinhaço Meridional (SdEM), na região sudeste do município de Diamantina, Minas Gerais. O clima da região é tipicamente subtropical de altitude, Cwb pela classificação de Koppen, caracterizado por verões brandos e úmidos e invernos mais frescos e secos (Sá Júnior et al., 2012).
Os quatro capões selecionados no PEB encontram-se entre as coordenadas 18°11’50,3”S 43°34’39,21”W e 18°11’20,37”S 43°34’34,6” W . Já os dezoito capões do PNSV estão localizadas na porção sul e norte, entre as coordenadas 17°55’40,7”S 43°49’48,8” W e 17°35'43.26"S 43°47'3.71"W (Figura 1).
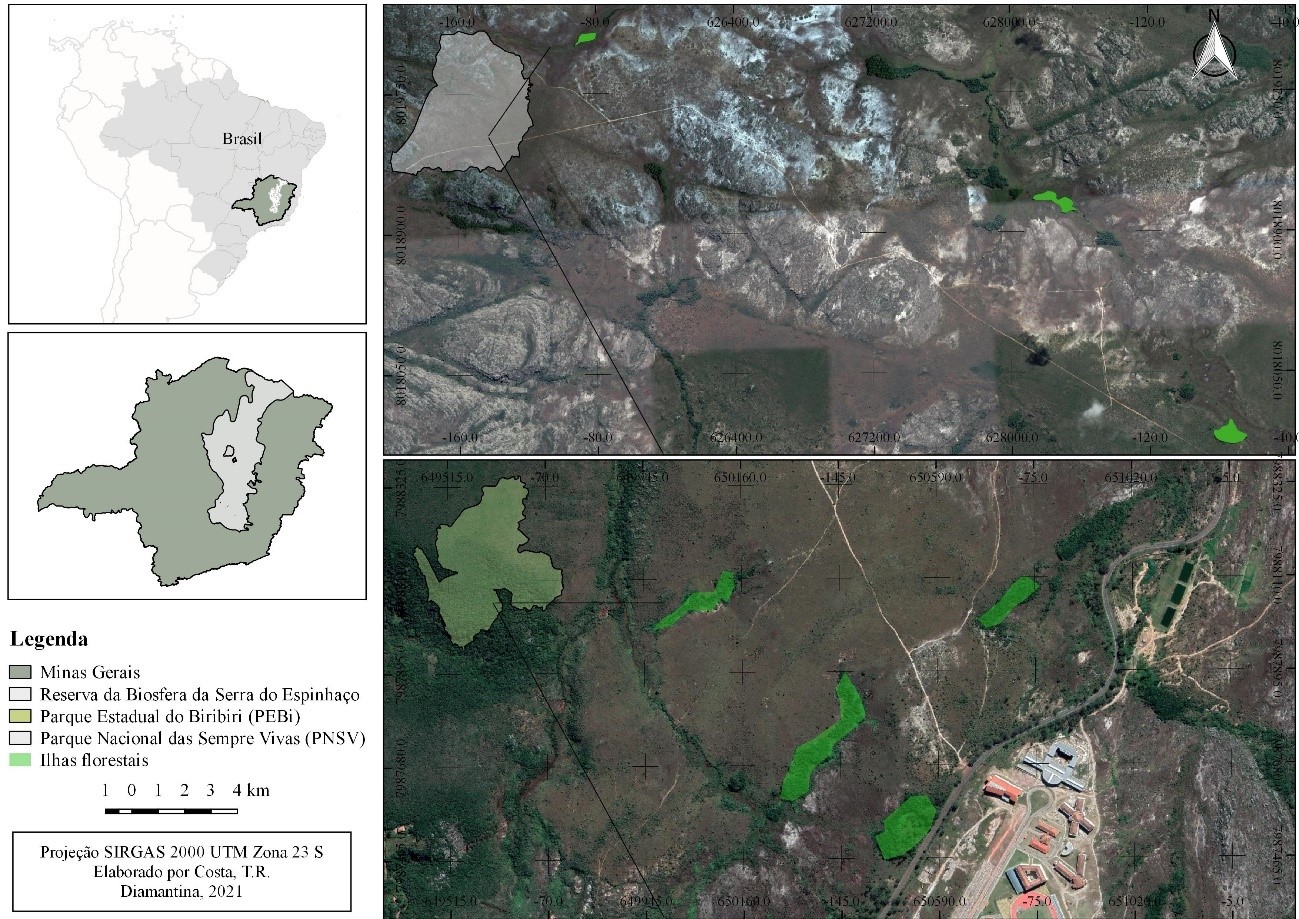 Figura 1. Localização dos
principais capões (ilhas florestais) amostrados no Parque Nacional das Sempre
Vivas (PNSV) e Parque Estadual do Biribiri (PEBi) – Reserva da Biosfera da
Serra do Espinhaço (RBSE)
Fonte: Google Satellite (2021).
Figura 1. Localização dos
principais capões (ilhas florestais) amostrados no Parque Nacional das Sempre
Vivas (PNSV) e Parque Estadual do Biribiri (PEBi) – Reserva da Biosfera da
Serra do Espinhaço (RBSE)
Fonte: Google Satellite (2021).
Todas as áreas amostradas estão distribuídas em altitude variando entre 1200 a 1380 m, distantes entre si, em no mínimo 200 m (Figura 1). Por estarem próximos ao topo das montanhas, estes ambientes estão dentro da zona de influência da Massa Atlântica Tropical oriunda da encosta leste (Domínio da Mata Atlântica) (Coelho et al., 2016). Ocorrem ao longo das nascentes dos cursos d’água e estão estabelecidas em manchas de Organossolos hidromórficos (Varajão et al., 2020), formando uma transição com as fitofisionomias secas adjacentes (Cerrado sentido restrito, campos rupestres e úmidos (i.e. “turfeiras”)). Assim, este sistema floresta-campo fornece importante serviço ecológico de proteção de regiões de nascentes e cabeceiras de afluentes de importantes bacias hidrográficas brasileiras (Coelho et al., 2018).
Os capões apresentam tamanho variável entre 0,02 e 3,0 ha, e se encontram em diferentes status de conservação, devido especialmente aos impactos causados pelo fogo (Figura 2). Cabe aqui a ressalva de que a prática da queimada ainda é muito comum no interior da área de proteção integral, para renovação de pastagens e coleta de sempre-vivas (Eriocaulaceae) (Sano e Giullieti, 2012). Sendo registrados no período de 2000 a 2014 aproximadamente 964 focos de incêndios dentro do território do PNSV e em suas proximidades (Soares, 2016).
 Figura 2 - Status
de conservação dos capões de matana Reserva da Biosfera da Serra do Espinhaço.
A: Ilha bem preservada, com alta densidade de epífitas. B: Ilha com intensa
abertura de clareiras devido à passagem do fogo.
Fonte: Costa (2017).
Figura 2 - Status
de conservação dos capões de matana Reserva da Biosfera da Serra do Espinhaço.
A: Ilha bem preservada, com alta densidade de epífitas. B: Ilha com intensa
abertura de clareiras devido à passagem do fogo.
Fonte: Costa (2017).
2.2 Composição
florística
Os capões foram inventariadas no mínimo 5 % de sua área total, por meio do sistema de amostragem aleatório. As parcelas permanentes tiveram dimensões de 10 x 10 m (100 m²), conforme sugerido pelo “Manual para o Monitoramento de Parcelas Permanentes nos Biomas Cerrado e Pantanal” (Felfilli et al., 2005). Assim, alocamos 72 parcelas nas ilhas florestais do PNSV, totalizando 0,72 ha de área amostral. Já no PEBi, o levantamento foi realizado em 12 parcelas (0,12 ha).
Foram amostrados todos os indivíduos com diâmetro à altura do peito maior ou igual a 3,0 cm (DAP ≥ 3,0 cm) obtidas a partir da circunferência à altura do peito (CAP). A circunferência foi medida com auxílio de fita métrica graduada em centímetros e a altura total com vara graduada em metros (projeção vertical do solo ao topo da copa). Não foram incluídas lianas e indivíduos mortos.
Foi realizada coleta do material botânico e posteriormente depositado no Herbário Dendrológico Jeanini Felfilli (HDJF), pertencente à Universidade Federal dos Vales do Jequitinhonha e Mucuri (UFVJM). A identificação das espécies, sempre que possível, foi realizada em campo, e também por meio de comparações em herbários, consulta a especialistas e bibliografia especializada.
As espécies encontradas nas ilhas florestais foram listadas em ordem de família e gênero. Para a classificação das famílias foi adotado o Angiosperm Phylogeny Group(APG) IV (2016) e os nomes de táxons foram atualizados usando o “International Plant Names Index-2012” (IPNI, 2012).
Foram realizadas consultas no Livro Vermelho da Flora do Brasil (Martinelli e Moraes, 2013) para averiguar quais espécies amostradas na área são consideradas ameaçadas de extinção. As categorias de ameaça foram definidas pela União Mundial para a Natureza (IUCN) e distribuem-se em: criticamente em perigo, em perigo, vulnerável, quase ameaçada, menos preocupante, dados insuficientes, extinta e extinta na natureza.
3. Resultados e
discussão
Para o conjunto total de capões, foram amostrados 1692 indivíduos totais, pertencentes a 40 famílias botânicas e 112 espécies (Tabela 1).
As famílias com maior
riqueza foram Myrtaceae (12), Fabaceae (10), Lauraceae (10), Rubiaceae (8) e
Melastomataceae (6). A maioria das espécies identificadas neste
estudo, ocorreram em outros estudos de matas de galeria inundáveis ( Silva, 2012; Gaya, 2014; Costa, 2017; Moura et al. 2021), e estão geralmente associados a solos com saturação
hídrica. Entre os trabalhos citados, a família Fabaceae é uma exceção, pois
possui pouca importância em estudos semelhantes, uma vez que, as espécies desta
família não se estabelecem plenamente em matas de galeria inundáveis (Costa,
2017), sendo a mesma, considerada a família mais representativa em termos de número
de indivíduos e de espécies em matas de galeria não inundáveis (Silveira, 2020)
Apesar das matas serem caracterizadas como inundáveis, algumas parcelas se encontravam mais drenadas em relação à maioria amostrada, e estas espécies ocorreram nestes sítios, especificamente. A ocorrência de espécies desta família botânica no presente estudo pode ter sido favorecida nestas parcelas em questão, próximas às bordas das matas, uma vez que, além de serem espécies generalistas de habitat, são indicadoras de locais mais abertos (Gonçalves et al., 2016). Estes resultados evidenciam que a maior heterogeneidade ambiental favorece a ocorrência de diferentes espécies, com requerimentos ambientais específicos, aumentando a riqueza em áreas com maior diversidade de ambientes (Silveira, 2020).
As espécies que mais se destacaram, em relação ao número de indivíduos foram: Richeria grandis ocupando o primeiro lugar no rank com 252 indivíduos, Xylopia emarginata (182 ind.), Protium spruceanum (149 ind.), Swartia multijuga (116 ind.) e Trembleyaa parviflora (102 ind.) (Tabela 1).
Moura et al. (2021) e Costa (2017) em seus estudos, apontaram a R. grandis como a espécie de maior valor de importância, corroborando para o resultado deste estudo. R. grandis apesar de ter uma especificidade ambiental única, apresenta uma distribuição geográfica ampla (Rabinowitz, 1981). Deste modo, é considerada uma das espécies mais representativas de ambientes ripários, caracterizados por solos hidromórficos e distróficos (Costa, 2017; Huber e Rull, 2019), uma vez que possui adaptações morfofisiológicas para suportar as condições físicas e a baixa oxigenação do solo nestes locais (Bambi et al., 2016), e é responsável por contribuir com mais de 45% da biomassa da serapilheira, determinante para a estabilidade destes ecossistemas (Rezende et al., 2017; Calderón et al. 2019).
Tabela
1. Florística dos 22 capões de mata amostrados no Parque Nacional das
Sempre Vivas (PNSV) e Parque Estadual do Biribiri (PEBi) – Reserva da Biosfera
da Serra do Espinhaço (RBSE).
 Fonte: Elaboração própria EM que: NI. número de indivíduos
Fonte: Elaboração própria EM que: NI. número de indivíduos
|
Família
|
Espécies
|
NI
|
PEBi
|
PNSV
|
|
Lamiaceae
|
Aegiphila integrifolia (Jacq.) Moldenke
|
1
|
x
| |
|
Euphorbiaceae
|
Alchornea glandulosa Poepp.
|
12
| |
x
|
|
Rubiaceae
|
Amaioua guianensis Aubl.
|
3
| |
x
|
|
Rubiaceae
|
Amaioua intermedia Mart.
|
1
| |
x
|
|
Fabaceae
|
Anadenanthera colubrina (Vell.) Brenan
|
3
| |
x
|
|
Fabaceae
|
Andira fraxinifolia Benth.
|
6
| |
x
|
|
Lauraceae
|
Aniba heringeri Vattimo-Gil
|
2
| |
x
|
|
Apocynaceae
|
Aspidosperma
cuspa (Kunth) S.F. Blake ex Pittier
|
6
| |
x
|
|
Asteraceae
|
Baccharis platypoda DC.
|
3
|
x
| |
|
Moraceae
|
Brosimum rubescens Taub.
|
26
| |
x
|
|
Malpighiaceae
|
Byrsonima sericea DC.
|
8
|
x
| |
|
Calophyllaceae
|
Calophyllum brasiliense Cambess.
|
70
|
x
|
x
|
|
Myrtaceae
|
Campomanesia velutina (Cambess.) O.Berg
|
1
|
x
| |
|
Salicaceae
|
Casearia grandiflora Cambess.
|
1
|
x
| |
|
Urticaceae
|
Cecropia pachystachya Trécul
|
24
|
x
|
x
|
|
Clethraceae
|
Clethra scabra Pers.
|
18
| |
x
|
|
Fabaceae
|
Copaifera langsdorffii Desf.
|
19
|
x
|
x
|
|
Rubiaceae
|
Cordiera macrophylla (K. Schum.) Kuntze
|
7
| |
x
|
|
Lauraceae
|
Cryptocarya moschata Nees & Mart.
|
1
|
x
| |
|
Sapindaceae
|
Cupania vernalis Cambess.
|
2
| |
x
|
|
Araliaceae
|
Dendropanax
cuneatus (DC.) Decne. & Planch.
|
6
| |
x
|
|
Winteraceae
|
Drimys brasiliensis Miers
|
12
| |
x
|
|
Lauraceae
|
Endlicheria
paniculata (Spreng.) J.F. Macbr.
|
2
| |
x
|
|
Asteraceae
|
Eremanthus incanus (Less.) Less.
|
25
|
x
|
x
|
|
Erythroxylaceae
|
Erythroxylum
pelleterianum A. St.-Hil.
|
9
| |
x
|
|
Erythroxylaceae
|
Erythroxylum sp.
|
1
|
x
| |
|
Myrtaceae
|
Eucaliptus sp.
|
3
|
x
| |
|
Myrtaceae
|
Eugenia florida DC.
|
3
|
x
|
x
|
|
Myrtaceae
|
Eugenia sp.
|
2
|
x
| |
|
Arecaceae
|
Euterpe edulis Mart.
|
37
| |
x
|
|
Rubiaceae
|
Faramea hyacinthina Mart.
|
19
|
x
|
x
|
|
Rubiaceae
|
Faramea nigrescens Mart.
|
4
| |
x
|
|
Rubiaceae
|
Ferdinandusa speciosa (Pohl) Pohl
|
19
|
x
|
x
|
|
Moraceae
|
Ficus insipida Willd.
|
2
|
x
| |
|
Moraceae
|
Ficus luschnathiana (Miq.) Miq.
|
4
| |
x
|
|
Arecaceae
|
Geonoma schottiana Mart.
|
1
| |
x
|
|
Myrtaceae
|
Gomidesia sellowiana O.Berg
|
4
|
x
| |
|
Nyctaginaceae
|
Guapira hirsuta (Choisy) Lundell
|
1
| |
x
|
|
Nyctaginaceae
|
Guapira opposita (Vell.) Reitz
|
6
|
x
|
x
|
|
Meliaceae
|
Guarea guidonia (L.) Sleumer
|
5
|
x
|
x
|
|
Bignoniaceae
|
Handroanthus
bureavii (Sandwith) S.Grose
|
1
|
x
| |
|
Bignoniaceae
|
Handroanthus ochraceus (Cham.) Mattos
|
1
| |
x
|
|
Bignoniaceae
|
Handroanthus serratifolius (Vahl) S.O.
Grose
|
1
| |
x
|
|
Chloranthaceae
|
Hedyosmum brasiliense Mart. ex Miq.
|
5
| |
x
|
|
Malpighiaceae
|
Heteropterys byrsonimifolia A. Juss.
|
1
| |
x
|
|
Aquifoliaceae
|
Ilex affinis Gardner
|
12
|
x
| |
|
Aquifoliaceae
|
Ilex cerasifolia Reissek.
|
1
|
x
| |
|
Fabaceae
|
Inga marginata Willd.
|
2
| |
x
|
|
Fabaceae
|
Inga nobilis Willd.
|
1
|
x
| |
|
Calophyllaceae
|
Kielmeyera lathrophyton Saddi
|
8
|
x
| |
|
Melastomataceae
|
Leandra scabra DC.
|
1
|
x
| |
|
Anacardiaceae
|
Lithraea molleoides (Vell.) Engl.
|
78
| |
x
|
|
Fabaceae
|
Machaerium punctatum (Poir.) Pers.
|
2
| |
x
|
|
Magnoliaceae
|
Magnolia
ovata (A.St.-Hil.) Spreng. |
2
| |
x
|
|
Euphorbiaceae
|
Maprounea guianensis Aubl.
|
1
| |
x
|
|
Myrtaceae
|
Marlierea
angustifolia (O.Berg) Mattos
|
1
|
x
| |
|
Celastraceae
|
Maytenus alaternoides Reissek
|
1
|
x
| |
|
Celastraceae
|
Maytenus sp.
|
3
| |
x
|
|
Melastomataceae
|
Miconia elegans Cogn.
|
17
|
x
|
x
|
|
Melastomataceae
|
Miconia ligustroides (DC.) Naudin
|
22
| |
x
|
|
Melastomataceae
|
Miconia theaezans (Bonpl.) Cogn.
|
19
|
x
|
x
|
|
Fabaceae
|
Moldenhawera emarginata (Spreng.)
L.P.Queiroz & Allkin
|
1
|
x
| |
|
Monimiaceae
|
Mollinedia schottiana (Spreng.) Perkins
|
4
| |
x
|
|
Celastraceae
|
Monteverdia robusta (Reissek) Biral
|
4
| |
x
|
|
Myrtaceae
|
Myrcia amazonica DC.
|
36
|
x
|
x
|
|
Myrtaceae
|
Myrcia deflexa (Poir.) DC.
|
10
| |
x
|
|
Myrtaceae
|
Myrcia pubescens DC.
|
10
| |
x
|
|
Myrtaceae
|
Myrcia splendens (Sw.) DC.
|
5
|
x
| |
|
Primulaceae
|
Myrsine
coriacea (Sw.) R.Br. ex Roem. & Schult.
|
7
|
x
|
x
|
|
Primulaceae
|
Myrsine gardneriana A. DC.
|
2
|
x
|
x
|
|
Primulaceae
|
Myrsine guianensis (Aubl.) Kuntze
|
3
|
x
| |
|
Primulaceae
|
Myrsine umbellata Mart.
|
3
| |
x
|
|
Lauraceae
|
Nectandra lanceolata Nees
|
11
|
x
|
x
|
|
Lauraceae
|
Nectandra megapotamica (Spreng.) Mez
|
7
|
x
|
x
|
|
Lauraceae
|
Nectandra oppositifolia Nees & Mart.
|
2
| |
x
|
|
Lauraceae
|
Nectandra warmingii Meisn.
|
1
| |
x
|
|
Lauraceae
|
Ocotea
aciphylla (Nees & Mart.) Mez
|
1
| |
x
|
|
Lauraceae
|
Ocotea corymbosa (Meisn.) Mez
|
2
|
x
|
x
|
|
Lauraceae
|
Ocotea puberula (Rich.) Nees
|
3
| |
x
|
|
Peraceae
|
Pera glabrata (Schott) Poepp. ex Baill.
|
2
|
x
| |
|
Asteraceae
|
Piptocarpha macropoda DC. Baker
|
1
|
x
| |
|
Melastomataceae
|
Pleroma candolleanum (Mart. ex DC.)
Triana
|
37
|
x
|
x
|
|
Rubiaceae
|
Posoqueria latifolia (Rudge) Schult.
|
21
| |
x
|
|
Burseraceae
|
Protium brasiliense (Spreng.) Engl.
|
1
|
x
| |
|
Burseraceae
|
Protium heptaphyllum (Aubl.) Marchand
|
40
|
x
|
x
|
|
Burseraceae
|
Protium spruceanum (Benth.) Engl.
|
149
| |
x
|
|
Malvaceae
|
Pseudobombax grandiflorum (Cav.)
A.Robyns
|
4
|
x
| |
|
Moraceae
|
Pseudolmedia laevigata Trécul |
11
| |
x
|
|
Myrtaceae
|
Psidium guineense Sw.
|
2
| |
x
|
|
Rubiaceae
|
Psychotria carthagenensis Jacq.
|
1
|
x
| |
|
Vochysiaceae
|
Qualea dichotoma (Mart.) Warm.
|
1
|
x
| |
|
Primulaceae
|
Rapanea umbellata (Mart.) Mez
|
6
|
x
| |
|
Phyllanthaceae
|
Richeria grandis Vahl
|
252
|
x
|
x
|
|
Proteaceae
|
Roupala montana Aubl.
|
1
|
x
| |
|
Araliaceae
|
Schefflera macrocarpa (Cham. &
Schltdl.) Frodin
|
14
|
x
| |
|
Araliaceae
|
Schefflera
morototoni (Aubl.) Maguire, Steyerm. & Frodin
|
10
| |
x
|
|
Anacardiaceae
|
Schinus terebinthifolia Raddi
|
1
| |
x
|
|
Fabaceae
|
Senna
reniformis (G.Don) H.S.Irwin & Barneby
|
2
|
x
| |
|
Simaroubaceae
|
Simarouba
versicolor A.St.-Hil.
|
1
|
x
| |
|
Myrtaceae
|
Siphoneugena densiflora O. Berg
|
14
| |
x
|
|
Fabaceae
|
Swartzia apetala Raddi
|
1
| |
x
|
|
Fabaceae
|
Swartzia multijuga Vogel
|
116
|
x
| |
|
Anacardiaceae
|
Tapirira guianensis Aubl.
|
23
|
x
|
x
|
|
Melastomataceae
|
Trembleya
parviflora (D. Don) Cogn.
|
102
|
x
|
x
|
|
Meliaceae
|
Trichilia hirta L.
|
1
|
x
| |
|
Hypericaceae
|
Vismia guianensis (Aubl.) Pers.
|
4
|
x
| |
|
Lamiaceae
|
Vitex polygama Cham.
|
2
|
x
|
x
|
|
Vochysiaceae
|
Vochysia thyrsoidea Pohl
|
6
| |
x
|
|
Vochysiaceae
|
Vochysia tucanorum Mart.
|
7
|
x
|
x
|
|
Annonaceae
|
Xylopia emarginata Mart.
|
182
|
x
|
x
|
|
Annonaceae
|
Xylopia
sericea A. St.-Hil.
|
5
| |
x
|
|
Salicaceae
|
Xylosma ciliatifolia (Clos) Eichler
|
1
|
x
| |
|
Total
| |
1692
|
62
|
75
|
As espécies Ferdinandusa speciosa, Geonoma schotiana, Cecropia pachystachya, Hedyosmum brasiliensis, além das supracitadas, são comumente abordadas na maioria das pesquisas desenvolvidas em ecossistemas de florestas ripárias (Gonzaga et al, 2019), onde figuram entre as mais abundantes. Dentre as espécies amostradas, algumas podem ser consideradas generalistas por compartilhar áreas de florestas e cerrado (Mendonça, 2008) como Copaifera langsdorffi, Maprounea guianensis, Trembleya parviflora, Tabebuia ocharea, Eremanthus incanus, dentre outras. Isto ressalta a influência das formações vegetais adjacentes na composição e diversidade destas florestas. Este resultado sugere que estes ambientes podem atuar como corredores ecológicos entre diferentes fitocenoses, e, portanto, pode constituir importante área de refúgio frente às futuras mudanças climáticas.
Das espécies amostradas, 50 ocorreram exclusivamente nos capões do PNSV, enquanto 37 espécies ocorreram apenas no PEBi. As duas áreas compartilharam apenas 25 espécies (22,3%) em comum (Figura 3). Este resultado sugere elevada diversidade beta entre as áreas, o que ressalta a importância singularidade e contribuição de cada Unidade de Conservação para a biodiversidade regional. Logo, estas áreas apresentam valor inestimável para manutenção dos processos de evolução (como especiação, fluxo gênico e adaptação) em zonas ecotonais (Souza, 2020).
O conhecimento da flora destes capõesnaturalmente isolados permite a extrapolação dessas informações para um plano geográfico, sendo uma ferramenta de grande valor prático para apoio à tomada de decisão. Com base nisso, evidenciamos a importância das seguintes medidas: i) manutenção e ampliação das Unidades de Conservação em locais com elevada diversidade ii) garantia da conectividade da paisagem por meio de corredores ecológicos (Morandi et al. 2020), visando favorecer o fluxo das espécies para diferentes fragmentos de floresta.
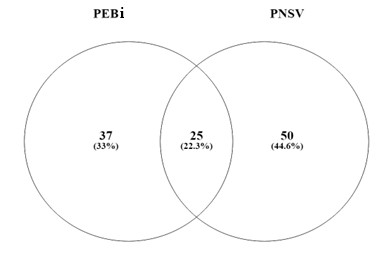 Figura 3. Diagrama de Venn
da composição florística dos capões do Parque Estadual do Biribiri (PEBi) e
Parque Nacional das Sempre- Vivas (PNSV), Reserva da Biosfera da Serra do
Espinhaço
Fonte: Elaboração própria
Figura 3. Diagrama de Venn
da composição florística dos capões do Parque Estadual do Biribiri (PEBi) e
Parque Nacional das Sempre- Vivas (PNSV), Reserva da Biosfera da Serra do
Espinhaço
Fonte: Elaboração própria
De acordo com Martins (1991), as espécies consideradas raras são aquelas que apresentam apenas um indivíduo na área amostrada. Com isso, as áreas deste estudo apresentaram 31 espécies raras, que representam 27,67 % das espécies amostradas e 1,18% dos indivíduos (Tabela 1).
Espécies que apresentam baixa densidade podem indicar duas situações distintas: i) que não tiveram muito sucesso na capacidade de colonização na comunidade, ou ii) pode tratar de um efeito da competição interespecífica dessas com as espécies já estabelecidas e melhor adaptadas às condições de alagamento do solo da área de estudo. Este padrão de ocorrência com muitas espécies representadas por poucos indivíduos apresenta fragilidade adicional dos ecossistemas fragmentados, pois a perda de alguns indivíduos pode ocasionar a perda de diversidade de espécies (Fagundes et al., 2019).
Dentre as espécies amostradas, quinze estão inseridas no Livro Vermelho da Flora do Brasil (2013), sendo estas: Euterpe edulis e Machaerium villosum categorizadas como vulneráveis e Nectandra warmingi, considerada como quase ameaçada. As espécies Andira fraxinifolia, Cletra scraba, Copaifera langsdorfii, Dendropanax cuneatum, Erythroxylum pelletelianum, Geonoma schotiana, Inga marginata, Maytenus robusta, Ocotea puberula e Mgnolia ovata estão inclusas na categoria de menos preocupante e Ocotea acyphila e Protium heptaphylum são consideradas espécies com deficiência de dados.
A composição de espécies categorizadas como ameaçadas de extinção e o elevado número de espécies consideradas raras acentuam, portanto, a importância da flora arbórea dos capões no Espinhaço.
4. Conclusão
Nossos
resultados demonstraram que capões naturalmente fragmentados da Reserva da
Biosfera da Serra do Espinhaço são ambientes que contribuem significativamente
para a biodiversidade regional, uma vez que constituem importantes centros
multitaxonômicos da flora. Adicionalmente, a identificação de espécies raras e
ameaçadas de extinção indicam a urgência de proteção efetiva destas florestas. Esperamos
que este trabalho possa nortear pesquisas futuras que garantam a conectividade
destas florestas, como métricas de ecologia da paisagem para implementação de
corredores ecológicos em Unidades de Conservação.
Agradecimentos
O
presente trabalho foi realizado com apoio da Coordenação de Aperfeiçoamento de
Pessoal de Nível Superior -Brasil (CAPES) - Código de Financiamento 001.
Referências
Alvarado S.T., Silva T.S.F., Fornazari et al. (2017). Drivers of fire occurrence in a mountainous Brazilian savanna: tracking long-term fire regimes using remote sensing. Ecological Indicators, 78, p. 270-281. https://doi.org/10.1016/j.ecolind.2017.02.037.
APG IV. (2016). An update of the Angiosperm Phylogeny Group classification for the orders and families of flowering plants: APG IV. Botanical Journal of the Linnean Society, 181, p. 1-20. https://doi.org/10.1111/boj.12385.
Arruda D.M., Schaefer C.E.G.R., Fonseca R.S. (2018). Vegetation cover of Brazil in the last 21 ka: new insights into the Amazonian refugia and Pleistocenic arc hypotheses. Global Ecology and Biogeography, 27, p.47–56. https://doi.org/10.1111/geb.12646
Bambi P., Rezende RS., Cruz T.M.S. et al. (2016). Diversidade da flora fanerogâmica de três matas de galeria no bioma cerrado. Heringeriana, v. 10, n. 2, p. 147-167.
Braga, S. S. et al. (2017) As chancelas da Unesco como alternativas de gestão para os patrimônios culturais e naturais da Serra do Espinhaço, Minas Gerais, Brasil. Revista Espinhaço, 1, p. 29-40. https://doi.org/10.5281/zenodo.3956622.
Bueno ML.; Pontara V., Neves DM., Ratter JA., Oliveira-Filho A.T. (2017). Effects of Quaternary climatic fluctuations on the distribution of Neotropical savanna tree species. Ecography, 40, n. 403-414. https://doi.org/10.1111/ecog.01860
Calderón C.D.C., Rezende R.S., Calor A.R. et al. (2019). Temporal dynamics of organic matter, hyphomycetes and invertebrate communities in a brazilian savanna stream. Community ecology, 20, p. 301-313. https://doi.org/10.1556/168.2019.20.3.10.
Coelho M.S., Fernandes G.W., Pacheco P., Diniz V., Meireles A., Santos R.M., Carvalho F.C., Negreiros D. (2016). Archipelago of montane forests surrounded by rupestrian grasslands: new insights and perspectives. In: FERNANDES GW. (EDS.) Ecology and Conservation of mountain-top grasslands in Brazil, Springer, New York, pp. 129-153.
Coelho M.S.; Neves F.S., Periilo L.N. et al. (2018). Forest archipelagos: A natural model of metacommunity under the threat of fire. Flora: Morphology, Distribution, Functional Ecology of Plants, v. 238, p. 244-249. http://dx.doi.org/10.1016/j.flora.2017.03.013
Costa T.R. (2017). Análise florístico-estrutural, relação vegetação-ambiente e transição floresta-campo das matas de galeria do Parque Nacional das Sempre-Vivas (PNSV), MG. 135 p. Dissertação (Mestrado) – Programa de Pós-Graduação em Ciência Florestal, Universidade Federal dos Vales do Jequitinhonha e Mucuri, Diamantina.
Fagundes N.C.A. (2019). Riparian vegetation structure and soil variables in Pandeiros river, Brazil. Rodriguesia, 70, p. 1- 19. http://dx.doi.org/10.1590/2175-7860201970002
Felfili J.M., Carvalho F.A.; Haidar R.F. (2005). Manual para monitoramento de parcelas permanentes nos biomas Cerrado e Pantanal. Brasília. Universidade de Brasília, Departamento de Engenharia Florestal. 55 p.
Fonseca B.M., Galvão L.M., Sousa FDR., Elmoor-Loureiro L.M.A., Souza MBG., Pinto R.L., Petracco P., Oliveira R.C., Lima E.J. (2018). Biodiversity in Pristine Wetlands of Central Brazil: a Multi-Taxonomic Approach. Wetlands, 38, p.145–156. https://doi.org/10.1007/s13157-017-0964-7
Fonseca SN. (2020). Efeitos do fogo em campos úmidos do Parque Nacional das Sempre-vivas, MG. 93p. Dissertação (Mestrado) – Programa de Pós-Graduação em Ciência Florestal, Universidade Federal dos Vales do Jequitinhonha e Mucuri, Diamantina, Minas Gerais.
Fontes CG.; Walter B.M.T. (2011). Dinâmica do componente arbóreo de uma mata de galeria inundável (Brasília, Distrito Federal) em um período de oito anos. Revista Brasileira de Botânica, 4(2), p.145-158. https://doi.org/10.1590/S0100-84042011000200002
Fontes C.G.; Walter B.M.T.; Pinto J.R.R.; Damasco G. (2015). Species turnover across different life stages for seedlings to canopy tress in Swamp Forests of Central Brazil. Internacional Journal of Ecology, 1. https://www.hindawi.com/journals/ijecol/2015/124851/.
Gaya T.R.L.M. (2014). A floresta inundável do norte de Minas Gerais: identidade florística e estrutura de comunidades arbustivo-arbóreas. 226 p. Dissertação (Mestrado) - Programa de Pós-Graduação em Ciências Florestais. Universidade Federal de Lavras, MG.
Gonçalves. S.T, Silva, R. H. R., Souza, S. R., Veloso, M. D. M., Nunes, Y. R. F. (2016) A vegetação dos afloramentos calcários na Serra do Cipó. Revista Espinhaço, 5, p. 19-31, doi: https://doi.org/10.5281/zenodo.3958074.
Gonçalves T.S., Silva A.C., Mendonça Filho C.V., Costa C.R., Braga I. L. (2020). The Capões of Seasonal Semi-deciduous Forest in the Cerrados and Rupestrian Fields of the Espinhaço Chain. International Journal of Geoscience Engineering and Technology, 1(1), p.43-48.
Gonzaga A.P.D., Machado E.L.M., Pinto J.R.R., Gripp A.M. (2019). Estudos florísticos e fitossociológicos em ambientes ripários das regiões Sudeste e Centro-Oeste do Brasil. Nativa, 7(5), p. 556-566, http://dx.doi.org/10.31413/nativa.v7i5.6817
Huber O., Rull V. (2019). Plant communities. Biodiversity of Pantepui: the pristine “lost world” of the neotropical Guiana Highlands, p.149-164.
IPNI (The International Plant Names Index). (2012). Disponível em: http://www.ipni.org/. Acesso em 10 de Julho de 2021.
Martinelli G., Moraes M.A. (2013). Livro Vermelho da Flora Brasileira. Andrea Jakobson Estúdio; Instituto de Pesquisas Jardim Botânico do Rio de Janeiro, Rio de Janeiro. 1100p.
Martins F.R. (1991). Estrutura de uma floresta mesófila. Campinas: UNICAMP, 245 p.
Mendonça RC., Felfili JM., Walter BMT., Silva Junior MC., Filgueiras TS., Nogueira PE., Fagg CW. (2008). Flora vascular do bioma Cerrado: checklist com 12.356 espécies. pp. 423-1279. In: Sano S.M., Almeida SP., Ribeiro JF. (eds.). Cerrado: ecologia e flora. v. 2. Brasília, Embrapa Informação e Tecnologia.
Mendonça-Galvão et al. (2011). Água do cerrado do Distrito Federal: biodiversidade, integridade e conservação. In: Fagg, C., Munhoz, C., Sousa Silva, J. Conservação de áreas de preservação permanente do cerrado: caracterização, educação ambiental e manejo. Brasília: CRAD, p.21-46.
Morandi D.T., Franca L.C.J., Menezes E.S., Machado E.L.M., Silva, M.D., Mucida D.P. (2020). Delimitation of ecological corridors between conservation units in the brazilian Cerrado using a GIS and AHP approach. Ecological Indicators, 115, p. 106440. https://doi.org/10.1016/j.ecolind.2020.106440
Moura C.C., Costa T.R., Oliveira P.A., Fonseca D.C., Machado E.L.M. (2021). Como é a estrutura e a diversidade alpha e beta de matas de galeria inundáveis? Diversitas Journal, 6(2), p.1920-1945. https://doi.org/10.17648/diversitas-journal-v6i2-1496
Oliveira M.C., Felfili J.M., Silva Júnior M.C. (2015). Comparação florístico-estrutural dos estratos adulto e de regeneração em Mata de Galeria perturbada no Distrito Federal, Brasil. R. bras. Bioci., Porto Alegre, 13(3), p. 142-154.
Queiroz, S. N.P., Pacheco, M. A. C. M., Santana, L. P., Cruz, C. C., Oliveira, C. N. S., Sincura, Y. R., Baracho, A. O., Souza, J. P., Rech, A. R. (2018). Polinizadores e visitantes florais da Cadeia do Espinhaço: o estado da arte. Revista Espinhaço, 7, p. 12-26. https://doi.org/10.5281/zenodo.3952802.
Rabinowitz D. (1981). Seven forms of rarity. In: Synge, H. (Ed.). The biological aspects of rare plant conservation, p205-217. New York: Wiley.
Rezende R.S., Sales M.A., Hurbath F. et al. (2017). Effect of plant richness on the dynamics of coarse particulate organic matter in a brazilian Savannah stream. Limnologica, 63, p.57-64. https://doi.org/10.1016/j.limno.2017.02.002
Ribeiro J.F., Walter B.M.T. (2008). As principais fitofisionomias do bioma Cerrado. In: Sano S.M., Almeida S.P., Ribeiro J.F. (Ed.). Cerrado: ecologia e flora. Brasília/DF: Embrapa Cerrados/Embrapa Informação Tecnológica, v.1, p.151-212.
Sá Júnior A., Carvalho L.G., Silva F.F., Alves M.C. (2012). Application of the Köppen classification for climatic zoning in the state of Minas Gerais, Brazil. Theoretical and Applied Climatology, 108(1), p. 1-7. https://doi.org/10.1007/s00704-011-0507-8
Sano PT., Giulietti AM. (2012). Eriocaulaceae. In: Wanderley M.G.L., Shepherd G.J., Melhem T.S., Guilietti A.M., Martins S.E. Flora fanerogâmica do estado de São Paulo. Instituto de Botânica, São Paulo. vol. 7, pp. 173-200.
Silva T.M.C., Carvalho W.A.C., Terra M.C.N.S. et al. (2016). Anthropic disturbances as the main driver of a semideciduous seasonal forest fragment in Minas Gerais. Rodriguésia, 70. https://doi.org/10.1590/2175-7860201970065
Silva W.M. (2012). Levantamento das espécies de dois trechos de mata de galeria na sub-bacia do Taquaruçu Grande, município de Palmas- TO: Composição, estrutura e relação planta-solo. 44p. Dissertação (Mestrado). Programa de Pós-graduação em Ecologia de Ecótonos da Fundação Universidade Federal de Tocantins.
Silva W.M., Lolis S.F., Viana R.H.O. (2016). Composition and structure of the gallery forest in the Taquaruçu Grande Sub-basin, Municipality of Palmas, Tocantins State. Acta Scientiarum. Biological Sciences, 38, n.1, p.17-24.
Silveira F.A.O., Negreiros N.P.U., Barbosa E. et al. (2016). Ecology and evolution of plant diversity in the endangered campo rupestre: a neglected conservation priority. Plant Soil, 403, p.129–152. https://doi.org/10.1007/s11104-015-2637-8
Silveira I.M. (2020). Padrões de ocorrência de espécies lenhosas em vertentes de Matas de galeria no Distrito Federal. 94p. Dissertação (Mestrado) – Programa de Pós-Graduação em Engenharia Florestal, Universidade de Brasília, DF.
Soares T.B.O. (2016). Avaliação de áreas queimadas no Parque Nacional das Sempre-Vivas – MG: Contribuições para a implantação do Manejo Integrado do Fogo. Dissertação (Mestrado) - Pós-Graduação em Geografia, UFMG, Belo Horizonte- MG. 124p.
Souza C.R.P., Paula G.G., Mendes C.N., Maia V.A., Aguiar-Campos N.C., Araújo F., Santos R.M. (2020). Local-scale tree community ecotones are distinct vegetation types instead of mixed ones: a case study from the Cerrado–Atlantic forest ecotonal region in Brazil. Australian Journal of Botany. pp 1-12 https://doi:10.1071/bt19108
Varajão, C. A. C., Varajão, A. F. D. C., Oliveira, F. S.; Barreto, H. N.; Salgado, A. A. R. (2020) Solos e superfícies de erosão: uma contextualização da evolução da paisagem na Serra do Espinhaço Meridional (SdEM), Minas Gerais. Revista Espinhaço, 9, p. 17-42. https://doi.org/10.5281/zenodo.3937475





