Centros: Revista Científica UniversitariaUniversidad de Panamá
EFECTO DE LA TEMPERATURA SOBRE EL CONSUMO DE OXÍGENO EN TILAPIAS DEL NILO CON DIFERENTES FENOTIPOS DE COMPORTAMIENTOEFFECT OF TEMPERATURE ON OXYGEN CONSUMPTION IN NILE TILAPIAS WITH
DIFFERENT BEHAVIOR PHENOTYPES
2021
102
04042021
28062021
HaniaNúñezhania.nunez@up.ac.paUniversidad de Panamá, Panama
ReynaldoVargasrvargas661@gmail.comUniversidad de Panamá, Panama
IvonneGuerraivonne.guerra@up.ac.paUniversidad de Panamá, Panama
EdgarRamosedgar800ramos@hotmail.comUniversidad de Panamá, Panama
Secciones
Referencias
Resumen
Servicios
Descargas
HTML
ePub
PDF
Buscar
Fuente
EFECTO DE LA TEMPERATURA SOBRE EL CONSUMO DE OXÍGENO EN TILAPIAS DEL NILO CON DIFERENTES FENOTIPOS DE COMPORTAMIENTO EFFECT OF TEMPERATURE ON OXYGEN CONSUMPTION IN NILE TILAPIAS WITH
DIFFERENT BEHAVIOR PHENOTYPES
Centros: Revista Científica Universitaria, vol.. 10, núm. 2, 2021 Universidad de Panamá
Resumen:
La tilapia del Nilo Oreochromis niloticus ha sido un pez introducido y adaptado a las condiciones tropicales para su producción acuícola. El objetivo de este estudio es determinar el efecto de la temperatura sobre el consumo de oxígeno en individuos separados por fenotipos de comportamiento. Los peces fueron separados en fenotipos de comportamiento utilizando la prueba de restricción y posteriormente sometidos a pruebas de consumo de oxígeno a temperatura ambiente e incrementando la temperatura utilizando una cámara respirométrica. Los resultados muestran que los peces proactivos asumen más riesgo para enfrentar un desafío en la prueba de restricción p<0.001. No se encontraron diferencias significativas en la tasa ventilatoria entre proactivos y reactivos p>0.05. Existe un efecto significativo en el consumo de oxígeno cuando se incrementa la temperatura p<0.001. Existe interacción entre la temperatura y el periodo de medición en el consumo de oxígeno en individuos proactivos p<0.001 y reactivos p<0.001. Estos resultados apoyan la hipótesis de que el fenotipo de comportamiento está asociada a el consumo de oxígeno en peces.
Palabras clave: Fenotipos de comportamientos, respirométria, cambio climático, Temperatura, acuicultura.
Abstract:
The Nile tilapia Oreochromis niloticus has been an introduced fish adapted to tropical conditions for its aquaculture production. The objective of this study is to determine the effect of temperature on oxygen consumption in individuals separated by behavioral phenotypes. The fish were separated into behavioral phenotypes using the restriction test and subsequently tested for oxygen consumption at room temperature and increasing the temperature using a respirometric chamber. The results show that proactive fish take more risk to face a challenge in the restriction test p <0.001. No significant differences were found in the ventilatory rate between proactive and reactive p> 0.05. There is a significant effect on oxygen consumption when the temperature increases p <0.001. There is an interaction between temperature and the measurement period in oxygen consumption in proactive individuals p <0.001 and reactive p <0.001. These results support the hypothesis that the behavioral phenotype is associated with oxygen consumption in fish.
La tilapia del Nilo se encuentra entre los peces de agua dulce más populares, que tiene un alto valor de mercado en el comercio mundial de pescado. Solo en China, el volumen de exportación de tilapia es de aproximadamente 1.100 millones. Actualmente China ha mostrado un incremento del cultivo de la tilapia del Nilo (Oreochromis niloticus) principalmente debido a los programas de mejora genética que han desarrollado muchas cepas adaptadas a diferentes condiciones climáticas, por ejemplo, la cepa GIFT (Tilapia de Cultivo Mejorada Genéticamente) que presenta ventajas, como la rápida tasa de crecimiento, el alto rendimiento de filete y la buena capacidad de resistencia a las enfermedades (Quiang et al., 2012).
O. niloticus
es un pez adaptado a las condiciones tropicales en sistemas de producción
extensivos e intensivos (Ono y Kubitza,
2003). Presenta tres características importantes para la acuicultura: la
primera es rusticidad es decir que puede producirse en todos los niveles de
intensificación (Fitzsimmons, 2000; Watanabe, Losordo y Fitzsimmons, 2002);
segunda, adaptabilidad a diferentes climas lo que permite su producción en
diferentes regiones del mundo (Avnimelech, 2007)
La calidad del agua está determinada por propiedades fisicoquímicas, una de las más importantes son la temperatura y el oxígeno. En tilapias del Nilo sus rangos de temperaturas oscilan entre los 25.0 y 32.0° C y su concentración de oxígeno disuelto es 5.0 - 9.0 mg/L; cuando se presentan cambios repentinos en la temperatura del agua el pez se estresa y este evento puede ocasionar su muerte (Saavedra-Martínez, 2006).
De acuerdo con Tomalá et al. (2014), en cultivos de peces, el oxígeno disuelto es el factor más importante de la calidad del agua, en bajas concentraciones puede causar pérdidas económicas significativas debido a sus efectos negativos en la ganancia de peso y representa la causa más frecuente de muerte de los organismos cultivados.
La temperatura a su vez afecta a la solubilidad del oxígeno disuelto en el agua, mostrando estos dos parámetros fisicoquímicos una relación inversa, es decir, a mayor temperatura menor solubilidad del oxígeno. En cuanto al consumo de oxígeno por los peces, resultados de investigaciones mostraron que hay una relación directamente proporcional, es decir, a mayor temperatura del agua mayor consumo de O2 disuelto (Valbuena- Villarreal y Cruz-Casallas, 2006).
Las diferencias individuales en las respuestas fisiológicas y de comportamiento a los estresores se han conservado evolutivamente y están presente en todos los taxones de vertebrados, incluidos los peces. Se consideran la materia prima para la evolución, por sus características adaptativas y pueden tener importantes implicaciones para los programas de selección de fenotipos que se desarrolla en acuicultura (Castanheira et al., 2015).
En los animales de granja, así como las especies explotadas en acuicultura, el interés en las asociaciones de rasgos consistentes de comportamiento y respuestas fisiológicas al estrés, es decir, los estilos de afrontamiento, ha aumentado dramáticamente en los últimos años, principalmente porque muchos estudios han demostrado que están vinculados con rasgos de desempeño, salud, resistencia a enfermedades y bienestar animal (Castanheira et al.,2013).
Estudios de perfiles genómicos señalan que los individuos de muchas especies de vertebrados muestran diferentes estilos de afrontamiento al estrés y estos tienen una influencia sobre cómo cambia la expresión génica en respuesta a una variedad de desafíos ambientales (Mackenzie et al. 2009).
En un estudio con lenguado senegalés (Solea senegalensis) se demostró que la selección por rasgos de comportamiento puede ser utilizada por la industria de la acuicultura para clasificar y seleccionar entre diferentes fenotipos mejorando los indicadores de producción (Zatarain et al., 2017).
En los estudios realizados por Martins et al. (2011), sobre el metabolismo basado en mediciones del consumo de oxígeno obtenido en cámaras de respirometría, los autores sugieren que la variación individual en consumo de oxígeno en una cámara de respirometría son relacionados a diferencias en estilos de afrontamiento.
Con estos antecedentes el objetivo de nuestra investigación fue determinar el efecto de la temperatura sobre el consumo de oxígeno en individuos separados por fenotipos de toma de riesgo.
Materiales y Métodos
Área de Estudio
Este
estudio se realizó durante el año 2019 en el laboratorio de acuicultura de la
Extensión Universitaria de Aguadulce del Centro Regional Universitario de Coclé
de la Universidad de Panamá, ubicada en el corregimiento El Cristo, distrito de
Aguadulce con coordenadas 545068.34 m E y 910037.96 m N.
Especie, población, alimentación y
aclimatación
Se utilizó alevines de tilapia del Nilo (Oreochromis niloticus) de aproximadamente 1 gramo. Los mismos se adquirieron en la estación experimental de la Autoridad de los Recursos Acuáticos de Panamá (ARAP), ubicada en Divisa, provincia de Herrera.
Un total aproximado de 300 individuos juveniles de ambos sexos formaron la población inicial. Los animales experimentales se trasladaron al laboratorio de acuicultura de la Extensión Universitaria de Aguadulce, fueron alojados en tanques de 400 litros equipados con un sistema de filtración mecánico y biológico. La densidad se estableció en 1.3 pez/Litro de agua. Se alimentaron hasta la saciedad dos veces al día durante el periodo de aclimatación con una tasa de alimentación del 5% de la biomasa. El periodo de aclimatación fue de un mes hasta que los animales no mostraron signos evidentes de estrés.
Parámetros físico-químicos del agua y fotoperiodo
Semanalmente se
recambió el 30% del volumen del tanque de alojamiento. La temperatura del agua
se mantuvo en 28.00±1.20ºC, las concentraciones de oxígeno se mantuvieron en
7.01±0.56 mg/L, adicionalmente se realizaron controles semanales de Amoniaco,
Nitrito, Nitrato y pH. El ciclo de luz/oscuridad se estableció en 12:12 (06:00
am-06:00 pm horas).
Diseño experimental
Se empleó un
diseño de tratamientos completos al azar con dos tratamientos
(proactivo-reactivo), se utilizaron 18 unidades experimentales; cada
tratamiento constó con 9 unidades experimentales. El modelo que se utilizó en
esta investigación fue el siguiente:
Yij= Son las variables respuestas de comportamiento (latencia), de estrés (respiraciones por minuto) consumo de oxígeno.
μ= es la media general;
Ei= es el efecto del fenotipo de comportamiento (i= proactivo, reactivo)
ei= es el error aleatorio.
Separación en fenotipos de comportamiento
Para la separación de los fenotipos de comportamiento se realizó
la prueba de restricción (Silva et al., 2010). La prueba de restricción
consistió en alojar cada pez en una malla fuera del agua por un minuto (Silva
et al., 2010). El aparato de prueba consistió en una malla para pecera de 16
centímetros de largo por 12 centímetros de ancho. La variable evaluada fue:
latencia primer escape.
Variable
Latencia primer
escape se definió como el tiempo tomado en segundos por cada pez para mostrar
un intento de escape. Un intento de escape se consideró cuando el pez levantó
su cuerpo de la malla (Castanheira et al., 2013).
Frecuencia Ventilatoria
La frecuencia
ventilatoria se midió en movimientos operculares por minuto (bpm - “beats per minute”
por sus siglas en inglés). Un movimiento opercular es igual a un ciclo de
apertura y cierre del opérculo (o boca) durante la respiración, es decir,
movimiento completo de inhalación y exhalación. Las observaciones se realizaron
visualmente desde uno de los laterales tapados, a través de una abertura (Calderer, 2001).
Criterios de selección entre fenotipos de comportamiento
Los criterios de selección empleado para la separación en fenotipos de toma de riesgo fueron establecidos en pruebas piloto preliminares. Individuos con latencias inferiores a 20 segundos fueron considerados proactivos, individuos con latencias entre 20 y 40 segundos fueron considerados intermedios y aquellos con latencias superiores a los 40 segundos fueron considerados reactivos. Los individuos intermedios fueron descartados en esta investigación.
Los peces de cada uno de los fenotipos de comportamiento fueron identificados empleando hilo de diferentes colores en la membrana ubicada entre la segunda y tercera espina de la aleta dorsal en individuos proactivos y entre la tercera y cuarta en individuos reactivos. Cada color correspondía a un individuo dentro del fenotipo de comportamiento.
Consumo de Oxígeno
El consumo de oxígeno disuelto de los fenotipos de comportamiento se evaluó con un modelo de cámara respirométrica adaptada de Tomalá et al., (2014). Se construyeron dos cámaras respirométricas con un volumen de 18 litros de agua, sellada con tapa hermética y una cubeta exterior de 14 litros de capacidad. Para ajustar la temperatura se utilizó un calentador. El registro del consumo de oxígeno se realizó con un multiparamétrico marca HACH modelo HQ40d con sonda para oxígeno. Adicionalmente se utilizó una bomba de agua.
Se introdujo la sonda de oxígeno en la cámara respirométrica, así como la bomba de recirculación de agua y se llenó el respirómetro con agua filtrada. Se suministró oxígeno procurando mantener el mayor porcentaje de saturación. La temperatura fue controlada con una cubeta de agua en el exterior del respirómetro y con un calentador sumergible dentro de una cubeta en el caso de incrementar la temperatura.
Determinación del consumo de oxígeno
Los peces fueron
introducidos individualmente en las cámaras respirométricas;
esto se llevó a cabo en dos fases distintas.
Los peces fueron introducidos individualmente en las cámaras respirométricas; esto se llevó a cabo en dos fases distintas.
Primera fase: Temperatura ambiente.
La primera fase se realizó a temperatura ambiente (± 28° C) donde se introdujo un pez proactivo y un pez reactivo en cada una de las cámaras respirometricas. Cada pez fue aclimatado durante 1 hora para reducir el estrés de manejo. El registro de consumo de oxígeno se realizó durante veinticinco minutos con registros a intervalos de 5 minutos.
Segunda fase: Incremento de temperatura.
En la segunda fase se realizó el incremento de 2 °C de la temperatura ambiente (30° C) introduciendo un pez proactivo y un pez reactivo en cada cámara respirometrica. Cada pez fue aclimatado durante 1 hora para reducir el estrés de manejo. El registro de consumo de oxígeno se realizó durante veinticinco minutos con registros a intervalos de 5 minutos.
Análisis estadístico
Todos los datos
fueron ingresados en una hoja de cálculo de Excel (Microsoft Excel
2018) para su procesamiento y el análisis estadístico se realizará con el
paquete estadístico Statistica versión 10
(StatSoftV10) y Graph Pad Prism V7 (San Diego, CA, USA). Las variables
respuestas evaluadas se examinaron por la prueba Levene de homogeneidad
de varianzas y la prueba de Shapiro-Wilk´s Test,
para verificar la distribución normal. Variables de la prueba de restricción y
tasa ventilatoria, peso y consumo de oxígeno se evaluaron utilizando t de studen´s o U de Mann-Whitney de acuerdo con los
preceptos de normalidad de homogeneidad de varianzas. Interacciones entre
fenotipo de comportamiento y tiempo de medida con Kruskal Wallis.
Resultados significativos en la prueba de Kruskal Wallis se evaluaron
con la prueba de comparaciones múltiples de Dunn’s.
Resultados
Prueba de Restricción
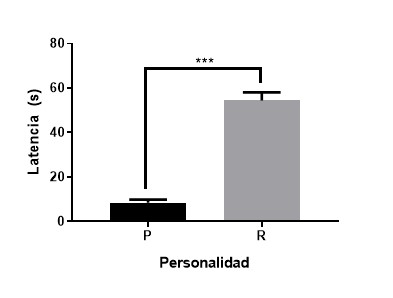
Los resultados muestran que los peces proactivos asumen más riesgo para enfrentar un desafío en la prueba de restricción. La prueba de Mann-Whitney muestra que existe diferencias significativas en latencia para realizar el primer intento de escape entre proactivos y reactivos (U= 0.00, p<0.001).
Ilustración
1 Media (± EE) de tasa
ventilatoria entre individuos proactivos (P) y reactivos (R). ***p>0.001.
Tasa
ventilatoria
Los
resultados muestran que no existen diferencias significativas entre peces
proactivos y reactivos en la tasa ventilatoria (t16=0.44, p= 0.66).
Peso
Los resultados muestran
que no existen diferencias significativas en el peso entre proactivos y reactivos
(t16=0.42, p= 0.67).
Temperatura
ambiente
Los resultados muestran
que no existe diferencia significativa en el consumo de oxígeno de forma
general entre proactivos y reactivos (U= 963.5, p=0.69). Los resultados
muestran que existe una tendencia en peces reactivos a consumir más oxígeno que
peces proactivos a los cinco minutos de medición (U= 19.5, p=0.06). Sin
embargo, no se mostró diferencia en el consumo de oxígeno entre proactivo y
reactivos a los 10, 15, 20, 25 minutos (U= 35.5, p=0.67; t16=1.55,
p= 0.14; U= 39.0, p=0.91; U= 38.5, p=0.88, respectivamente).
Incremento
de la temperatura
Los resultados muestran que proactivos y reactivos no mostraron
diferencias en el consumo de oxígeno a los 5, 10, 15, 20 y 25 minutos de
medición (U= 32.5, p=0.5; U= 37.5, p=0.81; U= 32.0, p=0.47; t16=1.21,
p= 0.24; t16=0.75, p= 0.46, respectivamente.
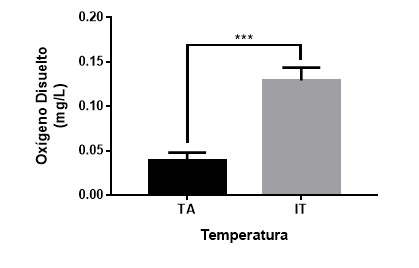
Temperatura
Los resultados muestran
que el incremento de la temperatura produce un aumento en el consumo de oxígeno
de los peces (U= 1161.0, p<0.001).
Ilustración
2 Media (± EE) de
consumo de oxígeno en temperatura ambiente (TA) y con incremento de la
Temperatura (IT). ***p>0.001.
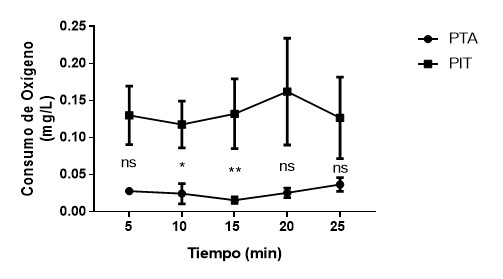
Proactivos
Los
resultados muestran que existe interacción entre la temperatura y el periodo de
medición en el consumo de oxígeno en individuos proactivos (K-W(9)= 48.43, p<0.001). La prueba de
comparaciones múltiples de Dunn´s indica que existe diferencias
significativas en el consumo de oxígeno entre proactivos en temperatura
ambiente y con incremento de la temperatura a los 10 y 15 minutos de medición
(p<0.05, p<0.01 respectivamente). Los tiempos 5, 20 y 25 minutos no
mostraron diferencias significativas (p>0.05).
Ilustración 3 Media (± EE) de peso entre individuos
proactivos en temperatura ambiente (PTA) y proactivos con incremento de la
temperatura (PIT). nsp>0.05,
*p<0.05, **p<0.01.
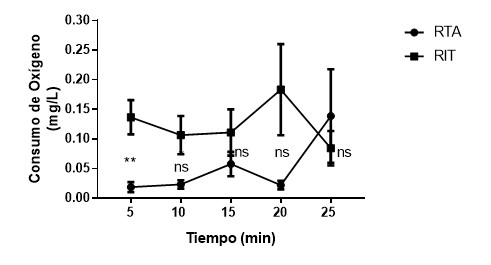
Reactivo
Consumo
de Oxígeno temperatura * periodo de medición
Los resultados muestran
que existe interacción entre la temperatura y el periodo de medición en el consumo
de oxígeno en individuos reactivos (K-W(9)=
33.22, p<0.001). La prueba de comparaciones múltiples de Dunn´s
indica que existe diferencias significativas en el consumo de oxígeno entre
reactivos en temperatura ambiente y con incremento de la temperatura a los 5
minutos de medición (p<0.01). Los tiempos 10,15, 20 y 25 minutos no
mostraron diferencias significativas (p>0.05).
Ilustración
4 Media (± EE) de peso
entre individuos proactivos en temperatura ambiente (PTA) y proactivos con
incremento de la temperatura (PIT). nsp>0.05,
**p<0.01.
Discusión
En este estudio se muestra con evidencias que la variación individual en respuestas de comportamiento en peces tiene importantes implicaciones en otros rasgos. Juveniles tilapias del Nilo mostraron variación individual en respuestas de comportamiento y en respuestas asociadas al metabolismo.
La prueba de restricción mostró ser una herramienta apropiada para evaluar la variación individual y agrupar los individuos en diferentes fenotipos de comportamientos como los descritos en la literatura (proactivo y reactivo). La latencia para realizar el primer intento de escape fue menor en individuos proactivos que reactivos en la presente investigación. Resultados similares se han reportado en doradas (Sparus aurata), en donde las latencias de individuos proactivos fueron más cortas que la de individuos reactivos, igualmente la prueba de restricción mostró consistencia temporal 14 días después de la primera separación por fenotipos de comportamiento. Además, el mismo estudio demostró que la prueba de restricción mostró consistencia contextual con las otras pruebas aplicadas (Castanheira et al., 2013).
Otro estudio realizado por Castanheira et al. (2015), mostró que los individuos proactivos mantenían latencias más cortas para realizar el primer intento de escape que individuos reactivos. En el mismo estudio se demostró que existen factores que pueden afectar la consistencia del fenotipo de comportamiento seleccionado por la prueba de restricción como puede ser el contexto social; estos investigadores mantuvieron grupos homogéneos de fenotipos de comportamiento y grupos con los fenotipos mezclados y encontraron que solo se observó consistencia en los grupos heterogéneos.
La tasa ventilatoria no fue predictiva de diferencias entre fenotipos de comportamiento en este estudio. Una posible explicación de este resultado puede ser la forma cómo se evaluó la variable, tomando el promedio de tres medidas repetidas realizadas sobre tres minutos; esto se apoya en el hecho que se considera que la tasa respiratoria es un indicador de estrés agudo (Barreto et al., 2003). También se ha reportado que la tasa ventilatoria en Gasterosteus aculeatus expuesto a un estresor mostró un rápido incremento, pero de corta duración (Bell et al. 2010). Además, Barreto y Volpato (2004), señalan que la tasa ventilatoria a pesar de ser un indicador sensitivo de respuesta de estrés no es proporcional al tiempo de exposición al estresor.
De acuerdo con los reportes de Falahatkar et al. (2019), en un estudio realizado con esturión belga (Huso huso), para evaluar el efecto de turbidez del agua por diferentes concentraciones de arcilla sobre la tasa ventilatoria se reportó una relación negativa entre tasa respiratoria y tiempo de exposición al contaminante. Otro dato interesante de esta investigación fue el hecho de que los grupos experimentales solo mostraron diferencias en la tasa ventilatoria durante el primer minuto de exposición, la diferencia desaparecía al segundo y tercer minuto.
En el presente trabajo realizado con juveniles de tilapia del Nilo se evaluó el efecto de la temperatura en el consumo de oxígeno en juveniles de tilapia del Nilo y se reportó un incremento del consumo de oxígeno asociado al incremento de la temperatura. Aumentos en el consumo de oxígeno asociadas a incrementos de la temperatura se han reportado en la literatura, por ejemplo, Cerezo y García (2004), reportaron incrementos en el consumo de oxígeno a medida que la temperatura se aumenta en la especie dentón común (Dentex dentex). En otro estudio realizado con tilapia roja (Oreochromis sp), se reportó que la temperatura tiene un efecto proporcional al consumo de oxígeno. Además, los autores explican que la temperatura dentro del medio acuático ocupa un lugar preponderante, ya que gobierna diferentes funciones del organismo y es uno de los factores que acelera las reacciones químicas y el metabolismo en general, ocasionando un mayor consumo de O2 por parte de los seres vivos (Valbuena-Villarreal y Cruz-Casallas, 2006).
En línea con estos resultados Galvis et al. (2005), en un estudio realizado con tilapia del Nilo (Oreochromis niloticus) con peso promedio de 86 gramos, comparando la alimentación a diferentes temperaturas, indicaron como resultado que de forma general la temperatura tuvo un efecto en el mayor consumo de oxígeno registrado por los peces. En nuestro caso las tilapias utilizadas mostraban menor peso que el estudio citado, sin embargo, los resultados fueron similares ya que el incremento de temperatura generó un incremento en el consumo de oxígeno de los peces experimentales.
También se evaluó si el fenotipo de comportamiento tiene un efecto en el consumo de oxígeno al incrementar la temperatura. Los resultados reflejan que el aumento de la temperatura produjo un incremento en el consumo de oxígeno de ambos fenotipos de toma de riesgo. Además, los resultados de este estudio reflejan respuestas diferenciales en el consumo de oxígeno durante el periodo de exposición en ambos fenotipos de comportamiento, por ejemplo, los individuos proactivos mostraron incrementos en el consumo de oxígeno al incrementar la temperatura a los 10 y 15 minutos de exposición, mientras que en reactivos el incremento se registró a los 5 minutos de exposición. Resultados de investigaciones relacionando a respuestas metabólicas asociadas al consumo de oxígeno y fenotipos de comportamiento se han evaluado en peces (Careu et al., 2008).
En estudio realizado por Huntingford et al. (2010), con carpa común (Cyprinus carpio), en individuos previamente separados por su disposición a tomar riesgo reportaron mayores tasas metabólicas en reposo asociadas al consumo de oxígeno en los individuos que asumían mayor riesgo. Igualmente, Jenjan et al. (2013), con Cyprinus carpio, reportaron que peces proactivos muestran mayores consumos de oxígeno que peces reactivos y argumentan que esta diferencia está asociada al mayor tamaño de estructuras relacionadas con el intercambio gaseoso en este fenotipo de comportamiento.
Por el contrario, resultados reportados por Martins et al. (2011), en Solea senegalensis, midiendo el consumo de oxígeno por 22 horas indican que se registró un menor consumo de oxígeno en individuos que mostraban comportamientos más rápidos al confinamiento, característico del fenotipo de comportamiento proactivos. En línea con estos reportes Herrera et al. (2014), en un estudio con Sparus aurata, reportaron un menor consumo de oxígeno en individuos que mostraban comportamientos característicos de evitar riesgo, es decir, de individuos reactivos.
En este estudio no se encontraron diferencias en el consumo de oxígeno como las reportadas por (Huntingford et al., 2010; Martins et al., 2011; Jenjan et al., 2013; Herrera et al., 2014) probablemente porque no se transformaron los datos de consumo de oxígeno a tasa metabólicas como en estos estudios citados; además se evaluó el consumo de oxígeno por un periodo de 25 minutos con muestreos cada 5 minutos y en la literatura citada existen diferentes tiempos de exposición y registro. Sin embargo, se encontró un efecto del fenotipo de comportamiento asociado al tiempo de respuesta en el consumo de oxígeno cuando la temperatura se incrementa.
Referencias Bibliográficas
Avnimelech, Y. (2007). Feeding with microbial flocs by tilapia in minimal discharge bio-flocs technology ponds. Aquaculture 264 (2007) 140-147
Barreto, R.E. y Volpato, G.L. (2004). Caution for using ventilatory frequency as an indicator of stress in fish. Behav. Process. 66 43–51
Barreto, R.E., Luchiari, A.C. y Marcondes, A.L. (2003). Ventilatory frequency indicates visual recognition of an allopatric predator in naïve Nile tilapia. Behav. Process. 60 235–239
Bell, A.M., Henderson, L. y Huntingford F.A. (2010). Behavioral and respiratory responses to stressors in multiple populations of three-spined sticklebacks that differ in predation pressure. J. Comp. Physiol. B – Biochem. Sys. Environ. Physiol. 180 211–220
Calderer, A. (2001). Influencia de la temperatura y la salinidad sobre el crecimiento y consumo de oxígeno de la dorada (Sparus aurata L.) (tesis doctoral) Universidad Autónoma de Barcelona, España.
Careau, V., Thomas, D., Humphries, M. y Reale, D. (2008). Energy metabolism and animal personality. The Authors. Journal compilation, 641-653.
Castanheira, M. F., Herrera, M., Costas, B., Conceicao, L. E. y Martins, C. (2013). Linking cortisol responsivess and aggressive behaviour in gilthead seabream Sparus aurata: Indication of divergent coping style. Applied Animal Behaviour Science. 143(2013) 75-81
Castanheira, M., Conceicao, L., Millot, S., Rey, S., Bégout, M., Damsgard, B., Kristiansen, T., Höglund, E., Øverli, Ø. y Martins, C. (2015). Coping styles in farmed fish: consequences for aquaculture. Reviews in Aquaculture 7, 1-19.
Cerezo, J. y Garcia, B. (2004) Influencia del peso y la temperatura sobre el consumo de oxígeno de rutina del Denton común (Dentex dentex Linnaeus,1758). Aguatic 21 16-23
Falahatkar, B., Bagheri, M. y Efatpanah, I. (2019) The efecto stocking on growth performance and biochemical índices in new hybrid of Leuciscus aspiusxRutilus frisii. Aquaculture Reports. 15 (2019) 1-7
Fitzimons, K. (2000). Tilapia: The most Important Aquaculture species of the 21 century. Eviromental Reserch Lab. University of Arizona.
Galvis, O., Cueto, A., Navarro y González, P. (2005). Consumo de oxígeno de juveniles de tilapia (Oreochromis niloticus) bajo diferentes condiciones de temperatura y su relación con la alimentación. Revista Dugandia, Ciencias Básicas, Uniatlántico, 1(1), 102-110.
Herrera, M., Castanheira, M. F., Conceicao, L. E., y Martins, C. (2014). Linking risk taking and the behavioral and metabolic responses to confinement stress in gilthead seabream Sparus aurata. Applied Animal Beahaviour Science 155 (2014) 101-108
Huntingford, F. A., Andrew, G., Mackenzie, S., Moreras, D., Coyle, S. M. y Kadri, S. (2010). Coping strategies in a strongly schooling fish, the common carp Cyprinus carpio. Journal of Fish Biology, 76, 1576-1591
Jenjan, H., Mesquita, F. Huntingford, F. y Adams, C. (2013). Respiratory function in common carp with different stress coping style: a hidden cost of personality traits? Animal Behaviour 1-5
Mackenzie, S., Ribas, L., Pilarczyk, M., Capdevila, D. M., Kadri, S., Huntingford, F. A. (2009). Screening for Coping Style Increases the Power of Gene Expression Studies. Plos One 4 (4) 1-5
Martins, C., Castanheira, M. F., Engrola, S.,
Costas, B. y Conceicao, L. E. (2011). Individual differences in metabolism predict coping
styles in fish.Applied Animal Behaviour
Science 130 (2011) 135-143
Ono, E.A. y Kubitza, F. (2003). Cultivo de peixes em tanques-rede. rev. e ampl. Jundiaí, 3-112.
Saavedra-Martínez, M. (2006). Manejo del cultivo de Tilapia. Managua, Nicaragua. USAID, University of Rhode Island, University of Hawaii and CIDEA, USA.
Silva P, Martins C, Engrola S, Marino G, Øverli Ø. y Conceição. (2010). Individual differences in cortisol levels and behaviour of Senega- lese sole (Solea senegalensis) juveniles: Evidence for coping styles. Applied Animal Behaviour Science, 124,75-81.
Tomalá, D., Chavarría, J. y Ángeles, B. (2014). Evaluación de la tasa de consumo de oxígeno de Colossoma macropomum en relación al peso corporal y temperatura del agua. Latin American Journal of Aquatic Research., 42(5), 971-979.
Valbuena-VillarreaL, R., y Cruz-Casallas, P. (2006). Efecto del peso corporal y temperatura del agua sobre el consumo de oxígeno de tilapia roja (Oreochromis sp). ORINOQUIA, 10(1), 57-63.
Watanabe, W.O., Losordo, T. M., Fitzimmons, K. y Hanley, F. (2002). Tilapia Production Systems in the Americas: Technological Advances, Trends, and Challenge. Reviews in fisheries Science, 10(3), 465-498
Zatarain, Z., Fatsini, E., Rey, S., Chereguini, O. Martin, I., Rasines, I., Alcaraz, C., y Duncan N. (2017). Royal Society Open Science 3, 2-18
Buscar:
Ir a la Página
IR
Modelo de publicación sin fines de lucro para conservar la naturaleza académica y abierta de la comunicación científica Visor de artículos científicos generados a partir de XML-JATS4R