
Secciones
Referencias
Resumen
Servicios
Descargas
HTML
ePub
PDF
Buscar
Fuente

REEVALUANDO LOS BOSQUES FÓSILES DE OCÚ
REVISITING THE OCU FOSSIL FORESTS
Tecnociencia, vol.. 21, núm. 1, 2019
Universidad de Panamá
Recepción: 27 Septiembre 2018
Aprobación: 30 Octubre 2018
Resumen:
RESUMEN
Las maderas fósiles de Ocú, conocidas como chumicos, han sido reconocidas por su abundancia y vistosidad, gracias a ello, han sido aprovechadas por los locales para su comercio. Los estudios realizados por Stern y Eyde en 1967 reportaron por primera vez las plantas fósiles de Ocú. Las familias y géneros reportados hasta el día de hoy parecen sugerir una vegetación relacionada con los bosques lluviosos. Esto ha sido confirmado en estudios previos y los primeros reportes de esta nueva investigación, en donde describimos diferentes maderas y un tallo con base en su anatomía interna, esto mediante la técnica de lámina delgada y su comparación taxonómica con maderas fósiles y actuales de distintas familias. Los resultados de las maderas estudiadas en este proyecto hasta ahora han sido identificadas a nivel de familia, en las que encontramos Arecaceae, Fabaceae, Moraceae, Anacardiaceae. Estos nuevos resultados referente a la anatomía como es la presencia de vasos mayores a 100 μm, ausencia de anillos de crecimientos, placas de perforación y presencia de palmas, confirman que desde hace 30 millones de años la paleoflora de la Península de Azuero era de bosques tropicales lluviosos.
Palabras clave: Maderas, fósiles, bosques, lluviosos, paleoflora, Azuero.
Abstract:
ABSTRACT
Fossil Woods from Ocu, locally known as chumicos, have been popular and highly commercialised due to their beautiful and attractive colours. Stern and Eyde (1967) published accounts on the Ocu fossil Woods for the first time. To date, taxonomic affinities on the reported taxa suggest rainforests dominated the Azuero region. These results have been confirmed in previous studies and with the first advances of this research. The fossil woods studied herein have been identified to the family level, namely, Arecaceae, Fabaceae, Moraceae and Anacardiaceae. First results confirm the dominance of rainforest taxa in Azuero.
Keywords: Woods, fossils, rainforests, paleoflora, Azuero.
INTRODUCCIÓN
El Istmo de Panamá, que ha constituido un puente de vida y al mismo tiempo, una barrera oceánica y biogeográfica por millones de años, ha sido poco explorado en temas paleontológicos, en particular con registros de plantas fósiles (Leigh et al., 2014; Bacon et al., 2015). Nuestro interés por la historia antigua del Istmo ha renacido tras nuevos hallazgos durante el proyecto de expansión del Canal de Panamá iniciado en 2007 (Herrera et al., 2014, MacFadden et al., 2014; Rodríguez-Reyes et al., 2014). Las formaciones geológicas en el Canal de Panamá no sólo han aportado importantes hallazgos de mamíferos fósiles, sino también sobre semillas y maderas, que evidencian relaciones con floras africanas, norteamericanas, pero especialmente suramericanas (Herrera et al., 2014; Rodríguez-Reyes et al., 2014). Entre las plantas encontradas en la Cuenca del Canal de Panamá, se han identificado maderas fósiles relacionadas con las familias Malvaceae (Guazumaoxylon miocenica, Periplanetoxylon panamense; Rodríguez-Reyes et al., 2014), Fabaceae (Prioria hodgesii; Rodríguez-Reyes et al., 2017.), Elaeocarpaceae (Panascleroticoxylon cristallosa; Rodríguez-Reyes et al., 2017), Chrysobalanaceae (Parinarioxylon; Jud et al., 2016) y otros árboles sólo identificados a nivel de familia como Melastomataceae,
Sapotaceae y Meliaceae.
A pesar de estos registros con plantas fósiles, Panamá tiene un alto valor paleontológico por descubrir en otras localidades que van más allá de la Cuenca del Canal, por ejemplo, la abundancia de maderas permineralizadas en el pueblo de Ocú dentro de la Península de Azuero es notoria y de gran potencial. Las maderas fósiles en Ocú y zonas aledañas, conocidas como chumicos, tienen tonos llamativos, atractivas para los lugareños que las han comercializado y utilizado con fines decorativos en comercios, casas y jardines (Fig. 1).

Sólo tres publicaciones formales abordan el tema de los árboles fósiles de Ocú, reportando la ocurrencia de Humiriaceae, Salicaceae y Hernandiaceae (Stern & Eyde, 1965); una nueva madera de Humiriaceae (Herrera et al., 2014); y recientemente Jud & Durnham (2017), reportan diez nuevos morfotipos relacionados con las familias Fabaceae, Lauraceae, Sapotaceae y Moraceae.
Las maderas fósiles se encuentran esparcidas en potreros y fincas, con poca evidencia de su roca parental, por lo mismo la edad de los bosques fósiles de Ocú permanece como un enigma. En un estudio previo realizado por Herrera et al. (2014) reportan limolitas en canteras en los alrededores de Ocú con fragmentos de madera carbonizada e invertebrados fósiles en sedimentos procedentes de la Formación Tonosí, datadas como del Eoceno, por lo que se sugiere que las maderas de Ocú están relacionadas con la Formación Tonosí. Por otra parte, Jud & Durnham (2017), mencionan que las maderas podrían relacionarse con la Formación Santiago, datada como del Oligoceno tardío a Mioceno temprano, sin embargo, aún sin evidencia de las maderas en su roca parental, lo que hace incierta su procedencia. Por otra parte, Jud & Durnham (2017) realizaron un Análisis de Escalamiento Multidimensional (NDMS), donde hacen una comparación de comunidades actuales usando caracteres anatómicos de la madera. Los resultados sugieren que los bosques fósiles de Ocú representan un antiguo Bosque Tropical Lluvioso, lo cual contrasta con los actuales bosques en la zona, bosques secos tropicales.
Los primeros resultados de nuestra investigación confirman los resultados previos de Jud & Durnham (2017) e incrementan el conocimiento de la composición florística de estos bosques fósiles.
MATERIALES Y MÉTODOS
Muestreo
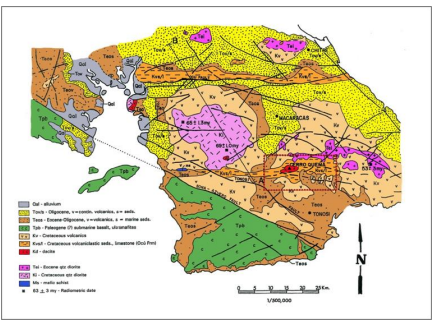
Se realizó una revisión de los mapas geológicos de Terry (1956), la Dirección General de Recursos Mineros (1976) (Fig. 2); Buchs et al. (2011) y Herrera et al. (2014) para ubicar las localidades con material fósil. Marcamos los puntos importantes sobre los mapas geológicos y establecimos los lugares a visitar. El estudio de los mapas nos ayudó a
identificar unidades geológicas en las localidades y a interpretar las secciones en los afloramientos; después nos comunicamos con los moradores del área y visitamos varias fincas privadas donde abundan las maderas permineralizadas de diversos tamaños.
Se realizaron dos visitas a campo con duración de una semana cada una. La primera salida de campo fue en mayo de 2017, donde se visitó varias fincas y potreros en las provincias de Herrera (Río Ocú, Los Pozos de Herrera y Valle Rico) y Veraguas (Llano de la Cruz) (); la segunda salida de campo realizada en mayo de 2018 incluyó las localidades en Los Pozos de Herrera y se adicionó una finca en Boquerones en Veraguas (Tabla 1, Fig. 3). En ambas visitas se recolectaron más de 100 maderas fósiles de diversos tamaños.
Los esfuerzos durante las primeras dos salidas de campo se concentraron en encontrar maderas in situ o identificar la roca parental de las maderas fósiles, ya que ha sido complicado determinar la
procedencia de estas abundantes y decorativas maderas fósiles. Tras el recorrido de varias fincas y la consulta a moradores conocedores del área, encontramos una sección de areniscas con trozos de madera fosilizada que corresponden con las maderas en estudio (Fig. 4). Destacamos que el hallazgo se atribuye a Miguel Martínez, joven estudiante de primer año de Biología en el Centro Regional Universitario de Herrera de la Universidad de Panamá.

Después de recorrer varias localidades en la Provincia de Herrera, el Sr. Carlos Sandoval un morador de Chitré, Herrera, dueño y coleccionista de maderas fósiles, nos hizo el favor de guiarnos a una de sus fincas, donde se encuentra el tronco fosilizado más largo hasta ahora encontrado en Panamá y posiblemente en todo Centro América (Fig. 5).

Preparación y análisis del material
Para la datación radiométrica de las rocas, las muestras de areniscas colectadas en Los Pozos y en Llano de la Cruz fueron etiquetadas y enviadas al Departamento de Ciencias Geológicas de la Universidad de Florida, Estados Unidos, para la extracción de zircones detríticos y su posterior análisis isotópico. Las muestras se encuentran bajo análisis al momento de este trabajo.
Para la parte taxonómica del proyecto, las maderas fósiles de gran tamaño se cortaron con un disco giratorio de punta de diamante para rocas en las instalaciones del Centro de Paleoecología y Arqueología Tropical (Center for Tropical Paleoecology and Archaeology). Esto se realizó con el fin de enviar piezas de tamaño manejable a Estados Unidos de América para la posterior preparación de secciones petrográficas. De las cuales, enviamos 33 de las muestras al Departamento de Biología de Adelphi University, Nueva York, Estados Unidos para la preparación de placas microscópicas o láminas delgadas. La técnica de lámina delgada consiste en obtener los cortes de los tres planos: corte transversal, tangencial y radial, para poder hacer la descripción completa para su posterior identificación taxonómica. De igual manera, enviamos 6 muestras a la Escuela Nacional de Ciencias Biológicas del Instituto Politécnico Nacional, México, donde el Dr. Emilio Estrada-Ruiz (Colaborador 2 de este proyecto) envió las muestras para la preparación de secciones petrográficas. Las secciones se prepararon siguiendo las técnicas de Jones & Rowe (1999). Los Drs. Estrada-Ruiz y Rodríguez-Reyes realizaron las descripciones e identificaciones de las muestras procesadas y fotografiadas en México, siguiendo las guías establecidas por la Asociación Internacional de Anatomistas de la Madera, *1989 (IAWA, por sus siglas en inglés), además se consultó bibliografía especializada sobre el tema (e.g., Metcalfe y Chalk, 1950; Détienne y Jacquet, 1983; Ilic, 1987, 1991; Terrazas, 1994), también nos apoyamos en InsideWood, una base de datos en línea para la identificación de maderas donde pudimos realizar una búsqueda más exhaustiva sobre las relaciones taxonómicas de nuestras maderas fósiles (InsideWood 2004-onwards; Wheeler, 2011).
Estos representan los primeros resultados del proyecto y nos encontramos escribiendo un artículo sobre la identificación del “Gran árbol” de Llano de La Cruz para ser enviado al Review of Palaeobotany and Palynology. Las muestras han sido identificadas a nivel de familia, remarcando algunos posibles géneros relacionados. Para nombrar nuevos géneros o especies fósiles, se requiere de un riguroso proceso de comparación en museos internacionales, por lo cual nos abstenemos de dar identificaciones a nivel género y especie hasta completar el proceso de comparación, que se llevará a cabo en un segundo año del proyecto.
RESULTADOS
Edad del material fósil
Durante las dos visitas a campo se corroboró la posición estratigráfica de las maderas fósiles de Ocú y, por lo tanto, inferir sobre su edad "tentativa" de ~30 Ma (Oligoceno) como parte de la Formación Santiago. Sin embargo, estas inferencias se basan en mapeos geológicos del área poco actualizados. La pobre preservación y afloramientos de los sedimentos donde se encuentran las maderas fósiles hacen difícil estudiarlas y conocer su procedencia geológica. Hemos realizado ensayos utilizando muestras de arenisca con trozos de madera y tovas volcánicas que subyacen la misma, aún sin poder corroborar su edad radiométrica o absoluta. Con lo cual se requieren repetir los análisis y sumar estudios bioestratigráficos con el fin de confirmar la edad del material.
Sistemática paleontológica
STRI ID: STRI 44038 B (medio) (Figura 4) “Gran árbol”
Familia: Anacardiaceae
Descripción:
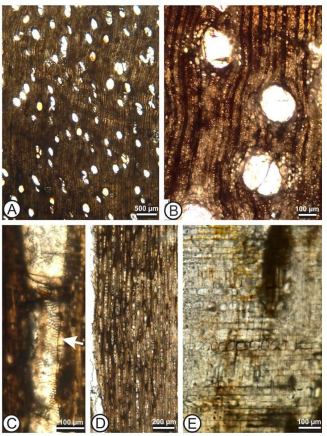
Madera con anillos de crecimiento indistintos y porosidad difusa; vasos solitarios combinados con unos pocos múltiplos radiales (34%) de 2–3 (–5); vasos con forma oval; placas de perforación simple; diámetro medio tangencial 154 (rango 96 – 205) μm; con 6 (rango 4 –
9) vasos por mm cuadrado; punteaduras intervasculares alternas, poligonales y medianas a largas (diámetros de 8 – 12 μm); punteaduras vaso-radio con bordes reducidos. Longitud de los elementos de vaso 365 (rango 203 – 504) μm. Tílides abundantes.
Parénquima axial paratraqueal, vasicéntrico y apotraqueal difuso y en algunos vasos con parénquima aliforme a confluente. Hebras de parénquima principalmente de 4-células. Radios heterocelulares mayormente uniseriados, con algunos biseriados. Radios compuestos de células mixtas en el cuerpo. Radios 19 (rango 15 – 23) por mm.
Fibras no septadas, con paredes de delgadas a intermedias 7.5 (rango 5
– 14) μm de grueso de pared y 33 (15 – 51) μm de lumen. Cristales
solitarios romboidales muy abundantes, especialmente abundantes en células erectas y rectangulares.
Comentarios:
Los caracteres presentes en la madera fósil de Ocú como porosidad difusa, vasos sin patrones de arreglo, vasos de tamaño pequeño a mediano (50 hasta <200 µm), placas de perforación simple, punteaduras intervasculares alternas y poligonales, tílides abundantes, fibras con paredes de grosor intermedio, parénquima paratraqueal y cristales prismáticos abundantes son caracteres comunes encontrados en Anacardiaceae (Terrazas, 1999). Después de revisar todas las especies disponibles en la base de datos de IWD (Insidewood-en adelante; Wheeler 2011) y literatura clave, encontramos varios caracteres presentes en el fósil que lo relacionan con el género Anacardium L. Entre los caracteres que encontramos similitud entre el fósil de Ocú y Anacardium son anillos indistintos, punteaduras intervasculares de tamaño grande (10 µm o >), punteaduras vaso-radio con bordes reducidos y parénquima paratraqueal vasicéntrico. Sin embargo, se requiere de comparaciones más exhaustivas y rigurosas con secciones micromorfológicas de todas las especies del género para sustentar dicha identificación.
Familia: Moraceae.
Descripción:
Madera con anillos de crecimiento indistintos y porosidad difusa. Vasos solitarios con unos pocos múltiplos radiales (32%) de 2 (–3); vasos de forma oval a redonda; placas de perforación simple; diámetro medio tangencial de vasos 270 (174 – 488 μm); media de 3 (rango 2 –
6) vasos por mm cuadrado; punteaduras intervasculares alternas, poligonales y grandes (diámetro de 11 a 15 μm); punteaduras vaso- radio con bordes reducidos, redondas a relativamente elongadas. Tílides abundantes.
Parénquima axial paratraqueal aliforme. Bandas marginales de parénquima, con hebras de 3 – 8-células. Radios 1 –3 células de ancho
y ocasionalmente de 4–10 células. Espaciamiento medio de radios 7 – 12 por mm linear.
Fibras septadas y no septadas con paredes de delgadas a gruesas, con 3 (1 – 4) μm de espesor de pared y 17 (12 – 23) μm de lumen.
Comentarios:
La presencia de caracteres como anillos de crecimiento indistintos, porosidad difusa, placa de perforación simple, punteaduras intervasculares alternas, poligonales y grandes, punteaduras vaso-radio con bordes reducidos, vasos con promedio de diámetro tangencial
>200 µm, tílides abundantes, fibras septadas y no septadas, parénquima apotraqueal aliforme y radios de 1-3 células de ancho. Los resultados sugirieron afinidades con Sextonia rubra (Lauraceae) y Prainea sp (Moraceae). Se descarta la similitud con Lauraceae, ya que no se encuentran células oleíferas en el fósil. Esta madera de Ocú tiene mayor afinidad con Moraceae, con la excepción de la tribu Ficeae. Caracteres como vasos >200 µm, presencia de fibras septadas y no septadas, punteaduras intervasculares > 10 µm, punteaduras vaso-radio con bordes reducidos, predominancia de parénquima apotraqueal y radios mayormente heterocelulares,
son características que distinguen a maderas de Moraceae (Welle et al., 1986; Ugwoke y Uju, 2010) y son caracteres que están presentes en el fósil aquí estudiado.
Familia: Leguminosae
Descripción:
Madera con anillos de crecimiento indistintos y porosidad difusa. Vasos solitarios combinados con 35% de múltiplos radiales de 2(–4); vasos en forma oval; Vasos con promedio de 6 (rango 3 – 10) vasos por mm cuadrado; placas de perforación simple; longitud promedio de elementos de vasos de 337 (rango 241 – 434) μm; punteaduras intervasculares alternas, diminutas (diámetro medio de 3 μm); punteaduras vaso-radio con bordes reducidos a aparentemente simples, redondas y grandes (rango 17 a 20 μm).
Parénquima axial paratraqueal vasicéntrico y parénquima apotraqueal difuso y con bandas de tres a cuatro células de grosor.
Radios de 1 a 3 células de ancho; radios multiseriados con células procumbentes y márgenes con tres filas de células cuadradas.

Cristales prismáticos presentes en las células cuadradas y parénquima axial.
Fibras no septadas, con paredes de grosor intermedio 2 (rango 4 – 1.5) μm de pared y 15 (rango 12 – 18) μm de lumen.
Comentarios:
Esta madera fósil presenta características propias de la familia Fabaceaae (conocidas también como Leguminosae) (Herendeen et al., 1996; Evans et al., 2006), los caracteres que sugieren afinidad de este fósil con las Leguminosae son la presencia de punteaduras intervasculares diminutas, parénquima apotraqueal abundante, fibras no septadas y radios heterocelulares. Sin embargo, para identificar el material a nivel de género e incluso de subfamilia, se requiere una comparación exhaustiva en las colecciones de micromorfología más completas del mundo.
Familia: Leguminosae Subfamilia: Detarioideae

Descripción:
Madera con anillos de crecimiento indistintos y porosidad difusa; vasos solitarios combinados con pocos múltiples radiales (33 %) de 2 (–3); vasos de forma oval; diámetro tangencial de 130 (rango 94 –150 μm).
Vasos de 6 (rango 4 – 9) vasos por mm cuadrado; placas de perforación simples; longitud de elementos de vaso 337 (rango 241–
434 μm); punteaduras intervasculares alternas, diminutas (diámetro medio de 3 μm); punteaduras vaso-radio similares a las punteaduras intervasculares. Fibras no septadas, con paredes muy gruesas.
Parénquima axial paratraqueal vasicéntrico.
Radios heterocelulares de 1 a 3 células de ancho; radios multiseriados con cuerpos procumbentes y 1- 2 filas de células marginales. Promedio de 11 radios por mm lineares.
Canales axiales normales presentes posiblemente presentes. Cristales abundantes.
Comentarios:
La presencia de canales axiales normales es un carácter relativamente inusual y solamente frecuente en las Dipterocarpaceae (Anisoptera, Dipterocarpus, Dryobalanus, Hopea, Shorea and Parashorea), Lauraceae (Cryptocarya), Rosaceae (Prunus), Vochysiaceae (Vochysia) y Leguminosae (subfamilia Detarioideae) (Rodriguez-Reyes et al., 2017). Al estudiar esta madera fósil, Dipterocarpaceae podría excluirse por poseer principalmente vasos solitarios y por tener radios anchos con 4–10 células. Las maderas de Cryptocarya, tienen punteaduras intervasculares grandes y células oleíferas asociadas al parénquima axial. Rosaceae (Prunus) posee anillos de crecimiento distintos, vasos en patrón diagonal y engrosamientos helicoidales en los vasos. Vochysia comúnmente tiene vasos grandes, parénquima abundante y canals de origen traumático. Las leguminosas de la subfamilia Detarioideae son las de mayor afinidad con esta madera fósil, basándonos en nuestra comparación.
Familia: Arecaceae
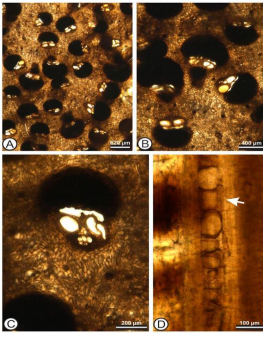
Descripción:
El cilindro central está delimitado por la zona cortical por un incremento de densidad, mientras el diámetro de fascículos vasculares reduce. No hay elongaciones de la zona fibro-cortical. El patrón general del tallo (clasificación Von Mohl; Von Mohl, 1845) es del tipo “Cocos”, con la densidad de fascículos fibro-vasculares siendo muy constantes.
La forma de la capa fibro-ventral es reniforme en corte transversal (clasificación Stenzel; Stenzel, 1904). No se observan sinus auriculares
en los fascículos vasculares. Los vasos del metaxilema son 3 (– 4) por fascículo fibro-vascular. Fitolitos globulares equinados. Parénquima basal del cilindro compacto. Parénquima basal del cilindro central compacto con células elongadas. Los fascículos fibro-vasculares tienen las siguientes dimensiones: Hmax (altura total de fascículos fibro- vasculares) Media= 626 μm, 557 – 880 μm; Hvasc (altura de la parte vascular) 260; 110 –140 μm. La media de diámetro de vasos=91. 14 μm (54 –188 μm). No se observa elongación de parénquima. 2 hebras de floema en la zona central.
Comentarios:
Este morfotipo de Arecaceae es diferente a los reportados de la paleoflora del Canal de Panamá, por tener 3 (– 4) vasos xilemáticos por fascículo fibro-vascular y parénquima basal del cilindro compacto con células elongadas. La mayoría de las palmas fósiles hasta ahora reportadas de Panamá parecen tener afinidad con las Coryphoideae, mientras que las características de este fósil corresponden con las Nypoideae.
DISCUSIÓN
Aunque se requiere extender los análisis de fechamiento o datación del material, para establecer una edad formal de los bosques fósiles de Ocú, logramos comprobar que la litología de las rocas relacionadas al “gran árbol” encontrado en Llano del Cruz (Finca del Sr. Sandoval) son las mismas que se encuentran en Los Pozos de Herrera, confirmando que son afloramientos de areniscas finas y lutitas de estratificación delgada.
Jud & Durnham (2017), describieron e identificaron 10 diferentes morfotipos de Ocú que revelaron afinidades con las familias Lauraceae, Arecaceae, Fabaceae, Moraceae, Euphorbiaceae, Humiriaceae, Anacardiaceae/Burseraceae y Simplocaceae. Además, aplicaron análisis paleoclimatológicos y por afinidades taxonómicas, concluyeron que los antiguos bosques fósiles de Ocú eran lluviosos, con un rango de precipitación (2000 mm por año) comparable a Barro Colorado y Panamá Central, contrastando con los rangos que
actualmente tiene la zona de Azuero, ~1000 mm al año. Comparamos los primeros resultados del proyecto con los de Jud y Durnham (2017) y menos de la mitad de los tipos de maderas coincide con las identificadas en los trabajos previos, por ejemplo, se han encontrado más especímenes de Andiroxylon barghornii y algunas Lauraceae similares a las previamente publicadas.
Todas las plantas identificadas de este estudio habitan bosques de Centro América (Gentry, 1982; Jaramillo et al., 2014) a excepción de Beilschmiedia que ocurre en rangos de bosques tropicales a templados. Las maderas descritas carecen de anillos de crecimiento y tienen porosidad difusa, sugiriendo muy poca estacionalidad y constante disponibilidad de agua. Estos resultados también están confirmados por el alto Indice de Vulnerabilidad obtenido (~2.1) y los resultados del Análisis de Escalamiento Multidimensional fuertes indicadores de bosques lluviosos tropicales (Jud y Durnham, 2017).
Jud y Durnham (2017) encontraron y colectaron especímenes de pequeño tamaño y sugirieron que los árboles de los Bosques fósiles de Ocú tenían alturas aproximadas de 21-27m. Sin embargo, de nuestro estudio, se han encontrado especímenes con >20 m de longitud y 2.5 m de perímetro preservado, sugiriendo una altura mínima de 35-40 m. Los árboles con mayores dimensiones hasta ahora encontrados en las localidades estudiadas corresponden con las familias Anacardiaceae y Moraceae.
El “gran árbol” (STRI 44038) presenta características de maderas de Anacardiaceae, en particular de Anacardium (espavé), un género netamente neotropical. Sin embargo, su registro fósil más antiguo se remonta del Eoceno de Alemania (Formación Messel) (Manchester et al., 2007), y se suman otros registros más recientes en América del Norte y Sur América. La historia paleogeográfica de Anacardium es compleja y sugiere una ruta biogeográfica compleja con algunos elementos que migraron desde Asia a Europa, pasando por América del Norte y llegando a Sur América; mientras que otros taxa se movieron desde el Sur Este de Asia la zona del Sub-Sahara y posteriormente a Sur América (Weeks et al., 2014). El espécimen de
Llano de La Cruz representaría el primer y más antiguo registro de este género en Centro América y es una pieza elemental para la reconstrucción biogeográfica de este género.
Dentro de las características de los bosques tropicales húmedos actuales es la gran diversidad de palmas (Kahn y de Granville, 1992) como lo atestigua el bosque fósil de Ocú. Durante este estudio, hemos encontrado al menos 4 morfotipos distintos de Arecaceae, no descritos antes para Panamá.
En el caso de Moraceae su distribución actual es en los bosques tropicales y subtropicales húmedos.
CONCLUSIONES
Los taxones identificados hasta ahora en nuestro proyecto confirman las conclusiones sobre el dominio de especies de bosques lluviosos en Azuero hace 30 millones de años aproximadamente. Estos resultados están confirmados también en el bajo porcentaje de anillos distintos en las maderas y estudios previamente realizados con Índice de Vulnerabilidad y Análisis de Escalamiento Multidimensional.
Las familias identificadas en este trabajo incluyen: Leguminosae, Moraceae, Lauraceae, Anacardiaceae, Arecaceae, Euphorbiaceae, Humiriaceae y Simplocaceae. Los árboles de mayores dimensiones encontrados de las colectas realizadas son de las familias Anacardiaceae y Moraceae.
Recomendamos dar continuidad a este proyecto y en su segunda fase, confirmar las identidades taxonómicas y sumar más morfotipos. También se requiere repetir el análisis radiométrico, utilizando las nuevas muestras recientemente colectadas en campo y realizar un estudio palinológico que permita obtener un rango de edad relativo.
Referencias
Bacon, C.D., D. Silvestro, C. Jaramillo, B. Tilston, Smith, P. Chakrabarty & A. 2015. Biological evidence supports an early and complex emergence of the Isthmus of Panama. PNAS: 201423853.
Buchs D. M., P.O. Baumgartner, C. Baumgartner-Mora, K. Flores & A.N. Bandini. 2011. Upper Cretaceous to Miocene tectonostratigraphy of the Azuero area (Panama) and the discontinuous accretion and subduction erosion along the Middle American margin. Tectonophysics. 512: 31–46.
Evans, J.A., P.E. Gasson & G.P. Lewis. 2006. Wood anatomy of the Mimosoideae (Leguminosae). IAWA J, Supp. 5.
Gentry, A.H. 1982. Patterns of Neotropical Plant Diversity. Evolut Biol. Pp 1 –84.
Herendeen, P.S., W.L. Crepet & D.L. Dilcher. 1992. The fossil history of the Leguminosae: phylogenetic and biogeographic implications. In: Herendeen, P.S., Dilcher, D.L. (Eds.), Advances in Legume Systematics, Part 4: The Fossil Record. Royal Botanic Gardens, Kew, pp. 303–316.
Herrera, F., S. Manchester, J. Vélez-Juarbe & C. Jaramillo. 2014. Phytogeographic history of the Humiriaceae (Part 2). Int J Plant Sci. DOI: 10.1086/676818.
International Association of Wood Anatomists Committee, 1989. List of Microscopic Features for Hardwood Identification. National Herbarium of the Netherlands. Leiden, NL. 332 pp.
Jaramillo, C.A., Moreno E, Ramirez, V.O., Silva-Caminha, S.A.F., de la Barrera A. 2014. Palynological record of the last 20 Million years in Panama. In: Paleobotany and Biogeography: A Festschrift for Alan Graham in his 80th year: 134–251. Miss Bot Gard Press, St. Louis, MO.
Jones, T. P. & N. Rowe. 1999. Fossil plants and spores: modern techniques. Geol Soc London.
Jud, N.A., C. Nelson & F Herrera. 2016. Fruits and wood of Parinari from the early Miocene of Panama and the fossil record of Chrysobalanaceae. Am J Bot 103 (2): 1 – 13.
Jud, N.A. & J. Durnham. 2017. Fossil woods from the Cenozoic of Panama (Azuero Peninsula) reveal an ancient neotropical rainforest. IAWA J 38 (3): 366-411.
Kahn, F. & J.J. de Granville. 1992. Palms in forest ecosystems of Amazonia. Berlín: Springer Verlag.
Leigh, E.G., A. O’Dea & G.J. Vermeij. 2014. Historical biogeography of the Isthmus of Panama. Biol Rev Cambridge Phil Soc. DOI: 10.1111/brv.12048.
MacFadden,B.J., J.I. Bloch, H. Evans, D.A. Foster, G.S. Morgan, A. Rincon, A.Wood. 2014. Temporal Calibration and Biochronology of the Centenario Fauna, Early Miocene of Panama The J of Geol 122:2, 113-135.
Manchester, S.R., V. Wilde & M.E. Collinson. 2007. Fossil Cashew Nuts from the Eocene of Europe: Biogeographic Links between Africa and South America. Int J Plant Sci 168 (8).
Rodríguez-Reyes, O., H. Falcon-Lang, M. Collinson, P.E. Gasson & C. Jaramillo. 2014. Fossil woods (Malvaceae) from the lower Miocene (early to mid-Burdigalian) part of the Cucaracha Formation of Panama (Central America) and their biogeographic implications. Review of Palaeobotany and Palynology 209: 11-34.
Rodríguez-Reyes, O., P.E. Gasson, H. Falcon-Lang & M.E. Collinson 2017. Two Panama Canal Miocene woods in the “Crudia” Group, Caesalpinioideae, Fabaceae. Review of Palaeobotany and Palynology. https://doi.org/10.1016/j.revpalbo.2017.06.005
Rodriguez-Reyes, O., P.E. Gasson, C. Thornton, H. Falcon-Lang & N.A. Jud. 2017. Panascleroticoxylon crystallosagen. et sp. nov: a new Miocene malpighialean tree from Panama. IAWA J 38 (4): 437-455.
Stern, W. L. & R. H. Eyde. 1963. Fossil Forests of Ocu, Panama. Science 1214.
Terrazas, T. 1999. Anatomía de la madera de Anacardiaceae con énfasis en los géneros americanos. Bol Soc Bot México 64: 103–109.
Terry RA. 1956. A geological reconnaissance of Panama. California Acad. of Sciences. 116 pp.
Ugwoke, C.E.C. & G.C. Uju. 2010. Anatomical studies of selected tree species of the Moraceae family grown in Nigeria. J pharmac all sci 7(1). http://dx.doi.org/10.4314/jophas.v7i1.63367.
Weeks, A., Zapata, F., Pell, S.K., Daly, D., Mitchell J.D., Fine, P.V.A. 2014. To move or to evolve: contrasting patterns of intercontinental connectivity and climatic niche evolution in “Terebinthaceae” (Anacardiaceae and Burseraceae). Front Genet 5:409. doi: 10.3389/fgene.2014.00409.
Welle, B.J.H., J. Koek-Noorman & S.M.C. Topper. 1986. The systematic wood anatomy of the Moraceae (Uriticales). V. Genera of the tribe Moreae without urticaceous stamens. IAWA J 7 (3).


