En el continente americano existen aproximadamente 10 especies de árboles de manglar (Field, 1995), estas especies pertenecen a las familias Avicenniaceae, Combretaceae, Pteridaceae y Rhizophoraceae. Algunas especies de mangle se pueden considerar especies ingenieras, ya que estabilizan el sustrato y áreas someras de las costas creando nuevos hábitats para otras especies animales y vegetales, si bien su crecimiento no se extiende mucho más allá de la línea de costa (Bossi y Cintron, 1989). De hecho, al ser un ecosistema dominado por especies leñosas, los manglares en Nicaragua representan una gran fuente de recursos económicos para las poblaciones locales donde las especies más afectadas por la tala son las especies del género Rhizophora.
Nicaragua por su posición geográfica posee manglares en sus dos costas, siendo siete especies arbóreas las que predominan: Laguncularia racemosa (L.) C.F. Gaerth. (mangle blanco), Avicennia germinans L., Avicennia bicolor Standl. (palo de sal), Conocarpus erectus L. (botoncillo), Rhizophora mangle L., Rhizophora harrisonii Leechm, y Rhizophora racemosa G. Mey, (mangle colorado)
A nivel mundial en el género Rhizophora se agrupan nueve especies (Tomlinson, 1999), este género está distribuido desde México hasta Ecuador, en el Caribe y en las costas de Brasil, en Nicaragua en la zona del Pacifico, podemos encontrar las especies R. mangle, R. racemosa y R. harrisonii, donde R. mangle domina en distribución ampliamente sobre las otras dos.
La descripción fenotípica generalizada para las especies R. mangle, R. harrisonnii y R. racemosa, nombrados comúnmente como mangles colorados, es que estas especies se caracterizan por ser árboles o arbustos de tierra litorales con numerosas raíces tipo aéreas, descendiendo de las ramas (Stevens et al., 2001). La altura de los árboles en este tipo de bosque oscila desde los 5-20 m de altura. Estas especies son vivíparas, lo que significa que la germinación ocurre cuando el embrión se encuentra todavía dentro del fruto y prendido del árbol progenitor (Sussex, 1975), llamándose propágulo al fruto con radícula. Rhizophora mangle, es común en pantanos costeros salobres y está distribuido en ambas costas (Stevens et al., 2001); Rhizophora harrisonii, esta especie es rara en manglares de la costa pacífica. La posición de R. harrisonii es morfológica y ecológicamente intermedia ente R. mangle y R. racemosa, lo que hace suponer su origen híbrido. Rhizophora racemosa no se ha diferenciado claramente de R. harrisonii; es una especie rara en manglares costeros, en sitios salobres. Rhizophora racemosa es una especie que la encontramos tierra adentro en los litorales del Pacífico de Nicaragua y Ecuador y desde el Caribe hasta Brasil (Stevens et al., 2001).
Las características morfológicas que permiten diferenciar a R. harrisonii de R. racemosa se asocian con la longitud y ancho de la inflorescencia, el número de flores, pero sobre todo por los diferentes números ramificaciones que presentan entre las tres especies. Según Jiménez (1994), estas características son elementos genéticamente muy plásticos. En el campo es posible distinguir inflorescencia y hojas con las características de estos tipos, pero es más frecuente aún encontrar inflorescencia y hojas con una marcada mezcla de estas características, incluso en un mismo árbol. La ausencia de una clara diferenciación en las características diagnósticas de estas especies hace muy difícil su separación en el campo.
Al encontrar que las tres especies del género Rhizophora presentan en principio características fenotípicas similares, las cuales hacen difícil su separación como especie, el objetivo propuesto para este trabajo es el de diferenciar morfofisiológicamente las tres especies del género Rhizophora, lo cual puede tener importancia para futuros diseños de estrategias de conservación del mangle colorado presente en las costas del Pacífico.
MATERIALES Y MÉTODOS
Material vegetal y lugar de estudio
El estudio se realizó con datos
fenotípicos y material vegetal del género Rhizophora
presente en la Reserva Isla Juan Venado (Nicaragua) ubicada a los 13º 66´ y
13º56´ de Latitud Norte y entre 49º 80´00” y 51º 33´ 00´´ de Longitud Oeste según
el Ministerio de Recursos Naturales y del Ambiente (MARENA et al., 2002)
(Figura 1).
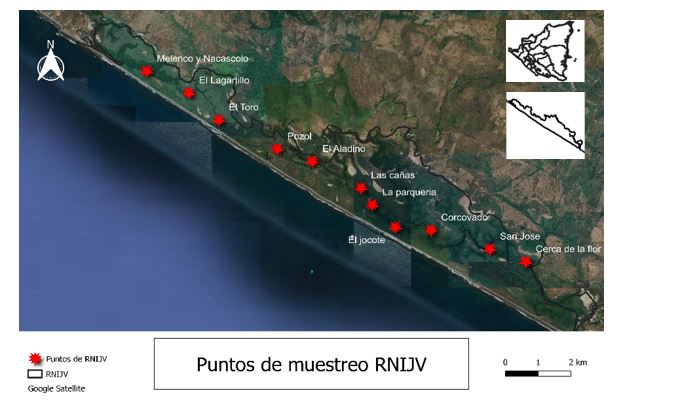 Figura
1.
Mapa de la Reserva Natural Isla Juan Venado (RNIJV). Los puntos rojos
señalan
los 11 puntos muestreados a lo largo de la isla. Autor: Carlos Solís (2022).
(Google Earth Satelite)
Figura
1.
Mapa de la Reserva Natural Isla Juan Venado (RNIJV). Los puntos rojos
señalan
los 11 puntos muestreados a lo largo de la isla. Autor: Carlos Solís (2022).
(Google Earth Satelite)
Diseño
experimental
El
diseño experimental fue el siguiente: se realizó una comparación morfológica de
las especies tomando datos morfológicos de 99 árboles muestreados a lo largo de
la isla, por otro lado, se hizo un recuento de lenticelas con muestras de 5
árboles por especies. La identificación
y diferenciación taxonómica de las especies se realizó mediante la revisión
bibliográfica de La Flora de Nicaragua de Stevens et al. (2001).
Morfología vegetal
Los datos fenotípicos se obtuvieron de 99 árboles en total a partir de los 11 puntos de muestreos que se realizaron a lo largo de la isla, se georreferenciaron los puntos muestreados (Figura 1); las características fenotípica tomadas fueron: altura total del árbol, altura de la raíz fúlcrea principal, grosor del tronco como diámetro a la altura del pecho (DAP), longitud total y anchura de la hoja, longitud del peciolo, número de lenticelas presente en la hoja, largo total de la inflorescencia, número de ramificaciones en la inflorescencia y longitud del pedúnculo floral.
El recuento de lenticelas se realizó en el envés de hojas de las tres especies. Debido a las grandes diferencias en densidad de lenticelas el sistema de contaje se adecuó a cada especie de la siguiente manera. Para R. mangle se contaron las lenticelas presentes. En una superficie de 1 cm2 en R. harrisonii y en R. racemosa en una superficie de 2 cm2. Para cada especie se realizó la densidad de lenticelas en cinco hojas.
Extracción
de pigmentos
La
extracción de pigmentos para las especies R. mangle, R. harrisonii
y R. racemosa se realizó a partir de material vegetal obtenido de las
hojas que previamente se había medido Fv/Fm. Los
discos foliares se seleccionaron al menos de 5 árboles de cada especie. Se
recolectaron teniendo presente tres variables; (i) de cualquier parte de la
hoja, (ii) con y (iii) sin
lenticelas. Los discos se conservaron con sílice gel para transportarlos al
laboratorio de Fisiología Vegetal de la Universidad del País Vasco según el
método de Esteban et al. (2009) para realizar la extracción de
pigmentos.
Análisis
de pigmentos
Los
espectros de absorción de cada pigmento se registraron de 250 a 700 nm. La
integración de los cromatogramas se realizó a 445 nm con el software Empower 2000. Los tiempos de retención y factores de
conversión son los mismos que en García-Plazaola y Becerril (2001). Este método
permite la separación de los pigmentos siguientes: neoxantina
(N), violaxantina (V), luteína epóxido (Lx), anteraxantina (A), luteína (L), zeaxantina (Z), clorofila b
(Chl b), clorofila a (Chl a), α-caroteno
(α-car) y b -caroteno (b-car). La separación de pigmentos
se realizó por HPLC con una columna de fase reversa C18 (Waters Spherisorb ODS1, 4,6 x 250 mm, Massachusetts, USA.), según
el método de García-Plazaola y Becerril (1999), con las modificaciones
descritas en García-Plazaola y Becerril (2001). La temperatura de la columna se
mantuvo a 35 ºC. Durante el proceso de HPLC las
muestras se mantuvieron a 4 °C en un
compartimento refrigerado. La identificación y la cuantificación se llevaron a
cabo con un detector de fotodiodos (PDA). Para cada muestra los pigmentos se eluyeron
siguiendo un siguiente gradiente de solventes (ver detalles en metodología).
Durante el procesamiento en el HPLC, las muestras se mantuvieron a 4 ºC en un compartimiento refrigerado.
Análisis de
tocoferoles
La detección de tocoferoles (Toc) se realizó con un detector de fluorescencia Waters 474. El método utilizado para la separación tocoferoles, la detección y cuantificación fue el mismo que en García-Plazaola y Becerril (1999 y 2001).
Los discos foliares se colocaron en tubos eppendorf de 2 ml, incorporando nitrógeno líquido para congelar completamente los tejidos y así impedir que la muestra vegetal se siga degradando, al mismo tiempo facilita la homogenización con el homogenizador Tearor 985370 (BioSpec, Bartslesville, USA) del tejido vegetal con acetona al 95 % (1500 µL). Se centrifugó a 13200 rpm durante 20 minutos a 4 °C, el sobrenadante se colocó en un nuevo tubo eppendorf y se filtró con filtros de 0.2 µm de PTFE (Teknokroma, Barcelona, España), colocándose en viales para su medida en el HPLC.
Estadística
Se
realizaron estadísticas con el programa SPSS versión 17.0 donde se realizaron
dos pruebas estadísticas: cálculo del valor de significación para las
correlaciones de los parámetros de caracterización fenotípica y morfológica; análisis
de varianzas (ANOVA) para la heterogeneidad de los pigmentos analizados de las
tres especies de Rhizophora, (p<0.05).
Antes del análisis ANOVA se comprobaron la homogeneidad de varianza con la
prueba de Levene y la normalidad de los datos con la prueba de Kolmogorov-Smirnov. Para los datos no paramétricos se
utilizó la prueba de Kruskal-Wallis.
RESULTADOS
Y DISCUSIÓN
Comparación
de la morfología de las tres especies
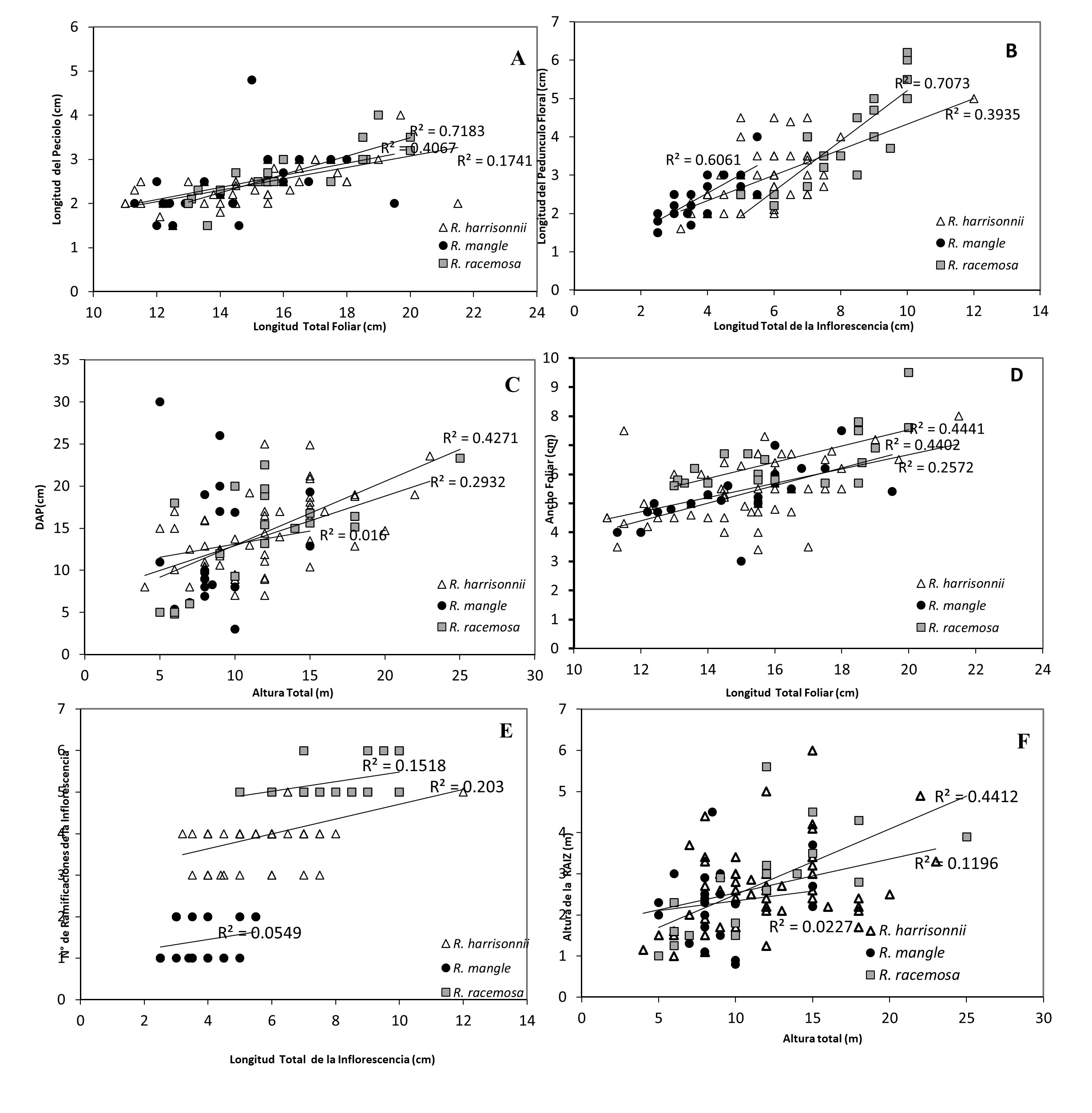
Como se muestra en la Figura 2 se puede observar que las características fenotípicas foliares son caracteres plásticos y que en si no diferencian las tres especies. Comparando la longitud total de las hojas para las tres especies (Figura 2 A, D), la máxima longitud es de 20 cm.
Las hojas presentaron un peciolo con longitudes variables que no las agrupan como especies. En cambio, teniendo en cuenta la característica de anchura de las hojas se puede decir que R. racemosa, seguido por R. mangle presentan hojas anchas, mientras que R. harrisonii puede tener hojas angostas o amplias como R. racemosa y R. mangle. Este hecho puede hacer que R. harrisonii sea clasificado erróneamente como una de las otras dos especies, cuando estos son clasificados por características fenotípicas de las hojas.
A diferencia de las hojas, una característica fenotípica que si se puede tomar en consideración para la clasificación de las tres especies es la inflorescencia. Ese es el caso de la longitud de pedúnculo floral (Figura 2B), R. mangle presentó una longitud menor que R. harrisonii en tanto que R. racemosa uno mayor. El número de ramificaciones (Figura 2E) presente en la inflorescencia también permitió la distinción entre especies, debido a que R. mangle presentó de 1-2 ramificaciones, R. harrisonii de 3-5 y R. racemosa de 5-6 ramificaciones, siendo esta característica de hecho, la principal característica fenotípica para diferenciar las tres especies.
La medida del Diámetro a la altura del pecho (DAP), es proporcional a la altura total del árbol (Figura 2C), donde se puede observar que cuanto más alto es el árbol presenta un mayor DAP. Lo mismo se puede atribuir a la altura de la raíz principal (Figura 2F) ya que por lo general cuanto más alto sea el árbol, la raíz tendrá una mayor longitud. En el caso de R. mangle estas relaciones no son tan claras, alcanzando además menor altura total que las otras dos especies.
Para la mayoría de los datos morfológicos se presentó una correlación positiva, siendo para la longitud del pedúnculo floral y el ancho de la hoja las características que mostraron diferencias significativas (p<0.05) entre las tres especies. La longitud del peciolo, el DAP y la longitud de la raíz principal, en cambio sólo presentaron diferencias significativas entre R. harrisonii y R. racemosa. Respecto al número de ramificaciones de la inflorescencia solamente R. harrisonii mostró diferencia significativa con el resto de las especies.
El género Rhizophora tiene un alto grado de variación poblacional como resultado de las variadas condiciones del ambiente tales como la concentración salina en el agua y las intensidades de irradiancia bajo las que las plantas de este género crecen. De acuerdo con Duke y Allen (2006), todas las especies del género Rhizophora tienen formas similares en cuanto a la morfología del árbol. Esta plasticidad morfológica que varía de acuerdo al ambiente en el que el árbol se desarrolle hace que no siempre se puedan distinguir entre las especies del género. Esto ha resultado en controversias sobre su clasificación taxonómica (Breteler, 1977).
 Figura
2.
Comparación fenotípica de caracteres morfológicos entre las especies R.
mangle, R. harrisonii y R. racemosa.
Longitud de la hoja (cm) (A) y Ancho de la hoja (cm) (D) en relación con el
largo total de la misma. Longitud del Pedúnculo (cm)
(B) y Número de ramificaciones de Inflorescencia (E)
comparados con el largo total de la inflorescencia (cm). Ancho del tronco (DAP)
(cm) (C) y Longitud de la raíz principal (m) (F) en comparación a la altura
total del árbol (m).
Figura
2.
Comparación fenotípica de caracteres morfológicos entre las especies R.
mangle, R. harrisonii y R. racemosa.
Longitud de la hoja (cm) (A) y Ancho de la hoja (cm) (D) en relación con el
largo total de la misma. Longitud del Pedúnculo (cm)
(B) y Número de ramificaciones de Inflorescencia (E)
comparados con el largo total de la inflorescencia (cm). Ancho del tronco (DAP)
(cm) (C) y Longitud de la raíz principal (m) (F) en comparación a la altura
total del árbol (m).
Las
tres especies de Rhizophora citadas para las
costas de América Latina (R. mangle, R. harrisonii
y R. racemosa), poseen características fenotípicas similares. Estas
especies pueden adaptarse y desarrollarse en una variedad de condiciones: desde
rocas hasta depósitos cenagosos y desde lugares con agua dulce, la mayor parte
del año, hasta en áreas donde la salinidad del suelo es superior a 21 g/L (Bossi
y Cintron, 1989; Lugo, 1981). Sin embargo, hay ciertas diferencias
ecológicas entre especies: R. mangle se desarrolla mejor en los suelos
poco profundos y cenagosos bajo la influencia de las mareas (Chapman, 1976), en
cambio R. harrisonii y R. racemosa crecen
en lugares que tengan proximidad a una fuente de agua dulce (Duke y Allen,
2006). Al comparar las características fenotípicas entre las tres especies,
unas de las características variables que podemos tomar en consideración y
relacionar con el ambiente en el que se desarrollan las especies del género Rhizophora es la de la altura total, que muestra que
los árboles con mayor altura son los de R. harrisonii
y R. racemosa. En cualquier caso, hay que aclarar que esto siempre va en
dependencia del lugar donde se desarrolle la especie, ya que por observaciones
de campo cuando éstas últimas especies se desarrollan en aquellos lugares donde
R. mangle compite y crece mejor no logran superar su altura.
Otra diferencia importante se observó en densidad de lenticelas en el envés de las hojas (Figura 3). En R. mangle el número de lenticelas por cm2 resultó ser el doble en comparación con R. harrisonii y R. racemosa, siendo este último el que presenta menor cantidad de lenticelas. Si bien no hubo diferencias significativas entre ambas especies (R. harrisonii y R. racemosa) (p>0.05).
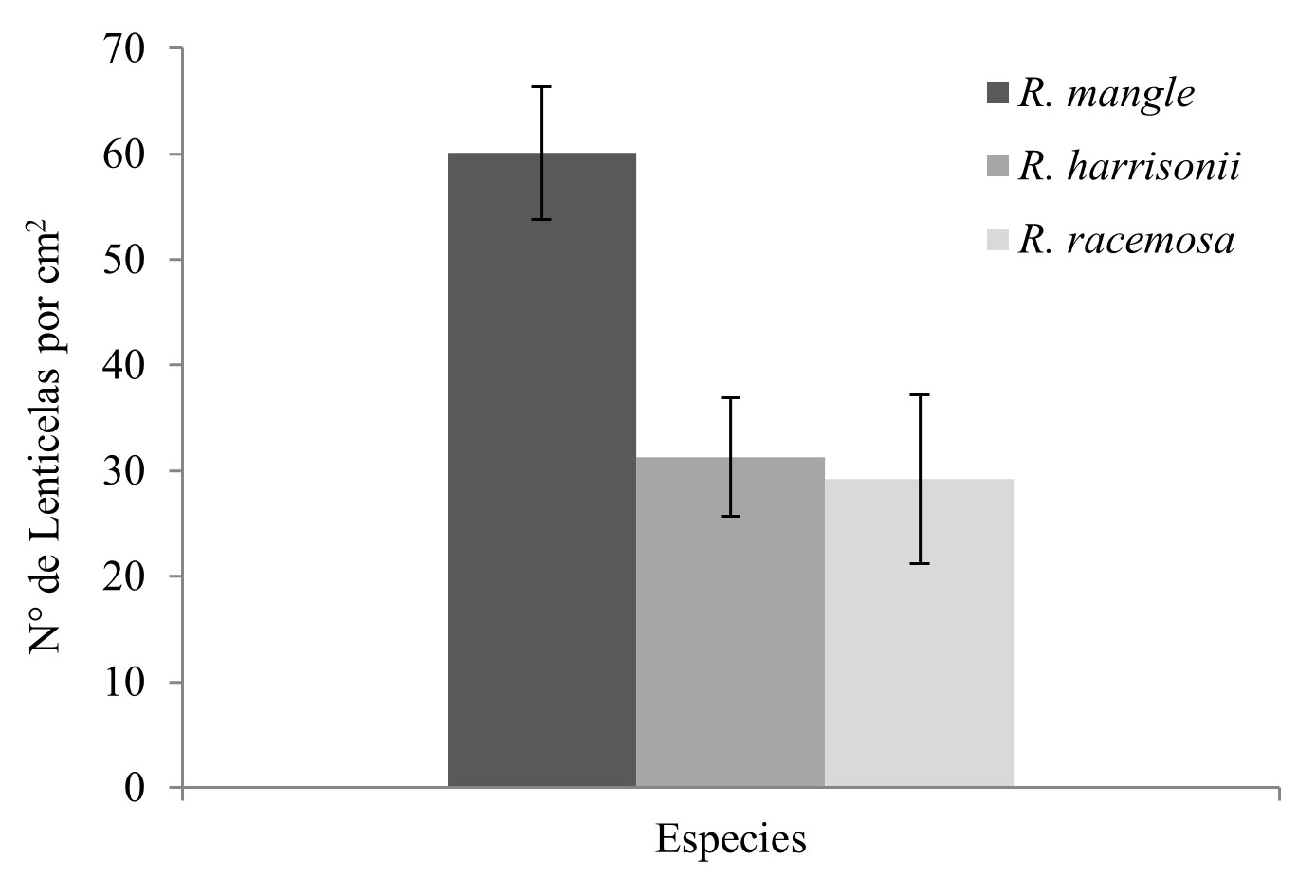 Figura 3.
Número de lenticelas por cm2
presentes en el envés de las hojas según especie.
Se muestran las
medias ± E.S (n≥5).
Figura 3.
Número de lenticelas por cm2
presentes en el envés de las hojas según especie.
Se muestran las
medias ± E.S (n≥5).
La cantidad de lenticelas también
varía incluso en el mismo árbol, presentando mayor cantidad de lenticelas las
hojas que están en contacto constante con el agua salada y con menor cantidad
las hojas que se encuentran en las partes más altas del árbol. Esta
distribución de las lenticelas es lógica considerando la función que ejercen
las lenticelas, ya que son estructuras permeables al aire, pero no al agua lo
cual facilita la respiración de las hojas que tienen contactos más frecuentes
con aguas salobres (Prahl, 1989).
Fotoprotección en las especies del género Rhizophora
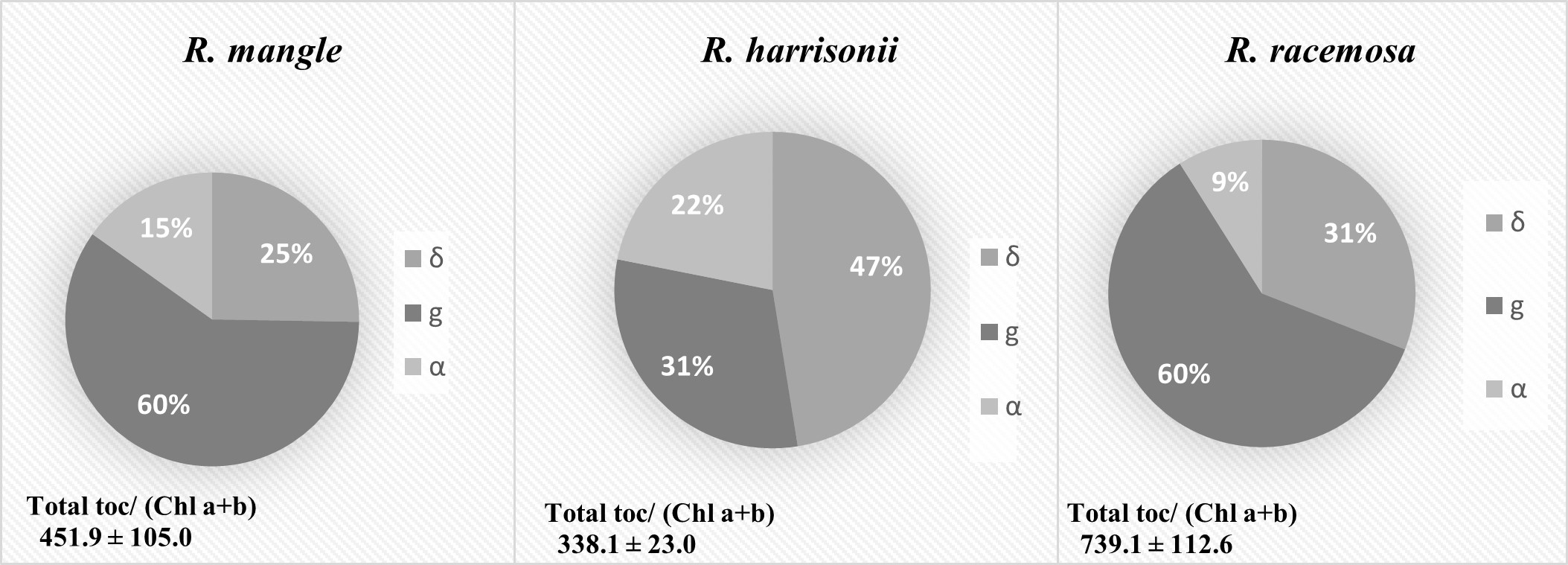 Figura 4.
Contenido total (mmol mol
Chl-1) y composición de tocoferoles (δ,g,α) en hojas para las
especies R. mangle, R. harrisonii y R.
racemosa. Se muestran valores medios ± ES. (n=5).
Figura 4.
Contenido total (mmol mol
Chl-1) y composición de tocoferoles (δ,g,α) en hojas para las
especies R. mangle, R. harrisonii y R.
racemosa. Se muestran valores medios ± ES. (n=5).
Los tocoferoles en las hojas de las tres especies de mangle mostraron diferencias tanto en el contenido como en la proporción entre los distintos tipos de tocoferoles. La especie que presentó mayor contenido del total de tocoferoles fue R. racemosa con 739.1 ± 112.6 (mmol mol Chl-1) seguida por R. mangle 451.8± 105.0 (mmol mol Chl-1), la Figura 4 nos muestra que para δ-toc, g-toc y α-toc las tres especies tienen diferentes proporciones. Para g- toc no hubo diferencia entre R.mangle y R. racemosa, en cambio para δ-toc y α-toc en las tres especies el contenido fue diferente.
El contenido de a-toc fue similar para las tres especies R. mangle 67 (mmol mol Chl-1), R. harrisonii 74 (mmol mol Chl-1) y R. racemosa 66 (mmol mol Chl-1). R. harrisonii al presentar mayor contenido de α-toc y menor contenido de g- toc en comparación a las otras dos especies, hace que R. harrisonii se diferencie de las otras dos especies a nivel bioquímico.
Además de tener diferentes contenidos en L y VAZ, la diferencia más notable entre las tres especies fue en el contenido de tocoferoles. Estas moléculas son antioxidantes lipofílicos que se sintetizan los organismos fotosintéticos y hongos (Sattler et al., 2006). Existen cuatro formas de tocoferol (α, b, g, y δ), de los cuales α-toc es la de mayor actividad antioxidante y la predominante en hojas (Munné-Bosch, 2005). El contenido de a-tocoferol aumenta bajo condiciones ambientales desfavorables (Chrost, et al., 1999), lo que indica su importancia fisiológica. En cooperación con otros antioxidantes a-tocoferol contribuye a la preservación de un adecuado estado redox en el cloroplasto durante la respuesta de las plantas al estrés (Munné-Bosch, 2005). Un número de estudios sugieren que la acumulación de a-tocoferol está relacionada con la tolerancia al estrés y una reducción está asociada con susceptibilidad al estrés (Leipner et al. 1999; Munné-Bosch y Alegre, 2000). Sin embargo, la caída de g-tocoferol metiltransferasa en plantas modificadas genéticamente (que cataliza la conversión de la g-tocoferol en el a-tocoferol), provoca una acumulación de g-tocoferol en lugar de a-tocoferol, y resulta en una mayor tolerancia a estrés osmótico y disminución de la tolerancia al estrés salino (Abbasi et al., 2007). En las tres especies de Rhizophora es llamativo su alto contenido de g-tocoferol que no suele estar presente en hojas, aunque si en semillas (Foryer y Noctor, 2003) (Figura 4). La acumulación de g-toc en lugar de a-toc en Rhizophora podría representar una respuesta al estrés salino. Pese a que el pool total de tocoferoles fue diferente en las tres especies, el contenido de a-toc fue muy similar. El a-toc es el tocoferol de mayor actividad antioxidante (Munné-Bosch, 2005), esto indica que las tres especies se encuentran sometidas a un nivel de estrés oxidativo similar. El contenido bajo de a-tocoferol en las tres especies también puede estar relacionado con la edad de la hoja, ya que el contenido de a-tocoferol es bajo en hojas jóvenes y aumenta con la edad de la hoja (Lichtenthaler, 1981; Szymánska y Kruk, 2008 a y b).
Al parecer R. harrisonii es la especie que tolera mejor el estrés por presentar menor contenido de tocoferoles en total y por presentar mayor porcentaje de a-tocoferol, mientras que la especie que se encuentra con mayor estrés es R. racemosa al no tener su entorno óptimo para su pleno desarrollo.
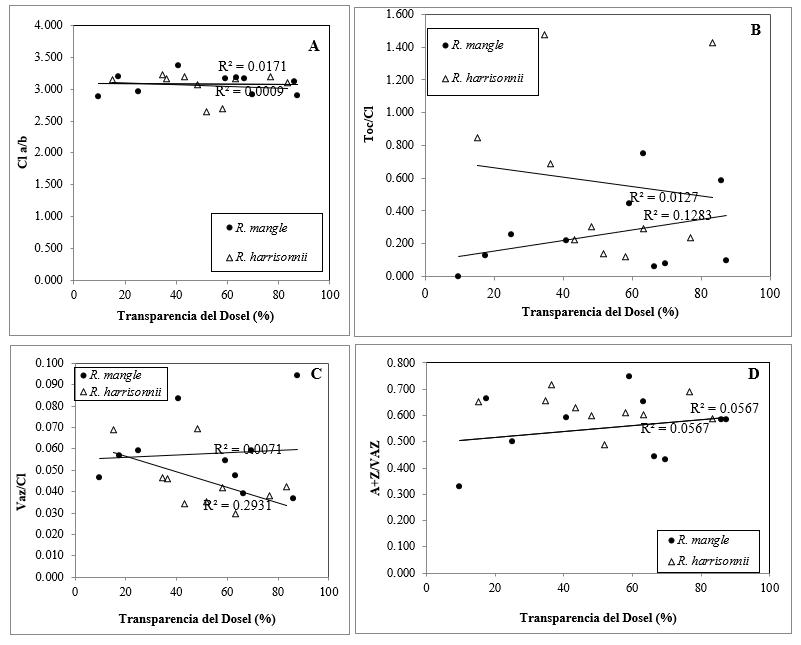 Figura 5.
Respuesta al porcentaje de transmitancia del dosel de los parámetros
indicativos de fotoprotección en hojas de Rhizophora mangle y R. harrisonii,
(n>5). Mol Clorofila a/mol Clorofila b (Chl a/b;
A); mol Tocoferol/mol Clorofila Toc/Chl;B); mol (Violaxantina+Anteraxantina+Zeaxantina)/mol Clorofila(a+b)(VAZ/Chl(a+b);C);
mol Anteraxantina+Zeaxantina/ mol Violaxantina+Anteraxantina+Zeaxantina
(A+Z/VAZ; D).
Figura 5.
Respuesta al porcentaje de transmitancia del dosel de los parámetros
indicativos de fotoprotección en hojas de Rhizophora mangle y R. harrisonii,
(n>5). Mol Clorofila a/mol Clorofila b (Chl a/b;
A); mol Tocoferol/mol Clorofila Toc/Chl;B); mol (Violaxantina+Anteraxantina+Zeaxantina)/mol Clorofila(a+b)(VAZ/Chl(a+b);C);
mol Anteraxantina+Zeaxantina/ mol Violaxantina+Anteraxantina+Zeaxantina
(A+Z/VAZ; D).
La
diferencia ecológica de las tres especies podría reflejarse en un diferente
nivel y demanda de fotoprotección. Sin embargo, las
diferencias fueron pequeñas y tan solo se observaron para los contenidos de
pigmentos fotosintéticos como L y VAZ (Figura 5). Tanto L como VAZ tienen
función fotoprotectora. Los pigmentos VAZ participan en el ciclo de las
xantofilas (Demmig-Adams y Adams, 1996) y la L está
involucrada en la estructura y organización complejas de la antena (Yan et
al., 2007) además tiene un papel fotoprotector (Dall´Osto et al., 2006; Yan et al., 2007) y una importante función como
antioxidante (Peng et al., 2006). Esto nos
indica que R. racemosa y R. mangle ocupan lugares expuestas a
mayor radiación solar en comparación a R. harrisonii.
Pese a las diferencias ecológicas entre las especies, las condiciones de
iluminación no parecen relacionarse con el potencial fotoprotector de cada
especie puesto que no se observó correlación significativa entre el porcentaje de la transparencia del dosel y el contenido en
pigmentos fotosintéticos de las especies de R. mangle y R. harrisonii (Figura 5). Esta poca plasticidad de los
sistemas fotoprotectores en relación con el ambiente lumínico, se puede atribuir a que aunque sean
diferentes especies y presenten diferentes ecologías en cuanto a su hábitat,
estas son especies que se adaptan a cualquier medio de las otras especies del
género Rhizophora, adoptando así
características fenotípicas similares y con ello la fotoprotección
puede deberse más a características morfológicas que a la cantidad de contenido de pigmentos fotosintéticos.
CONCLUSIONES
Las características morfológicas de las hojas y la presencia de lenticelas no son fiables para la diferenciación de las especies del género Rhizophora del Pacífico de Nicaragua, ya que estas características son adaptables al medio donde el árbol se desarrolle.
Rhizophora harrisonii presentó menor contenido de ganma tocoferol (g-toc) lo que podría relacionarse con su peor tolerancia al estrés salino. Además, pese a existir diferencias en el contenido total de tocoferol, la cantidad de alfa-tocoferol (α-toc) fue similar en las tres especies, lo que indica que están sometidas a un nivel similar de estrés oxidativo.
El contenido de tocoferoles puede ser una herramienta para la clasificación de estas especies principalmente para separar a R. harrisonii y R. racemosa como especies. Aunque estas dos especies son las que presentan mayor similitud en cuanto a las características morfológicas e incluso en las ramificaciones de las inflorescencias, su distinta composición en tocoferoles podría convertirse como parámetro bioquímico de distinción interespecífica.
REFERENCIAS BIBLIOGRÁFICAS
Abbasi, A. R., Hajirezaei, M., Hofius, D., Sonnewald, U., & Voll, L. M. (2007). Specific roles of α- and g- tocopherol in abiotic stress responses of transgenic tobacco. Plant Physiology, 143(4),1720–1738. https://doi.org/10.1104%2Fpp.106.094771
Bossi, R. y Cintron, G. (1989). Manglares del Gran Caribe. Hacia un Manejo Sostenible. Asociación para la conservación del Caribe.
Breteler, F. J. (1977). America’s Pacific species of Rhizophora. Acta Botanica Neerlandica, 26(3), 225-230
Chapman, V. J. (1976). Mangrove vegetation. J. Cramer.
Chrost, B., Falk, J., Kernebeck, B., Molleken, H., & Krupinska K. (1999). Tocopherol biosynthesis in senescing chloroplasts—a mechanism to protect envelope membranes against oxidative stress and a prerequisite for lipid remobilization?. En J. H. Argyroudi-Akoyunoglou & H. Senger (Eds.), The chloroplast: from molecular biology to biotechnology (pp. 171–176). Kluwer Academic Press.
Dall´ Osto, L., Lico, C., Alric, J., Giuliano, G., Havaux, M., & Bassi, R. (2006). Lutein is needed for efficient chlorophyll triplet quenching in the major LHCII antenna complex of higher plants and effective photoprotection in vivo under strong light. BMC Plant Biology, 6. https://doi.org/10.1186/1471-2229-6-32
Demmig-Adams, B., & Adams, W. W. (1996). The role of xanthophylls cycle carotenoids in the protection of photosynthesis. Trends in Plant Science, 1(1), 21-26. https://doi.org/10.1016/S1360-1385(96)80019-7
Duke, N. C., & Allen, J. A. (2006). Rhizophora mangle, R. samoensis, R. racemosa, R. × harrisonii (Atlantic-East Pacific red mangroves). https://www.doc-developpement-durable.org/file/Culture/Arbres-Bois-de-Rapport-Reforestation/FICHES_ARBRES/Arbres-non-classes/Rhizophora-AEP.pdf
Esteban, R., Baloguer, L., Manrique, E., Rubio de Casas, R., Ochoa, R., Fleck, I., Pintó-Marijuan, M., Casals, I., Morales, D., Jiménez, M. S., Lorenzo, R., Artetxe, U., Becerril, J. M., & García-Plazaola, J. I. (2009). Alternative methods for Sampling and Preservation of photosynthetic pigments and tocopherols in plant material from remote locations. Photosynth Res, (101), 77-88. https://doi.org/10.1007/s11120-009-9468-5
Field, C. D. (1995). Journey Amongst Mangroves. International Society for Mangrove Ecosystem.
Foryer, C. H., & Noctor, G. (2003). Redox sensing and signaling associated with reactive oxygen in chloroplasts, peroxisomes, and mitochondria. Plant Physiology, 119(3), 355–364. https://doi.org/10.1034/j.1399-3054.2003.00223.x
García-Plazaola J.I., & Becerril J.M. (2001). Seasonal changes in photosynthetic pigments and antioxidants in beech (Fagus sylvatica) in a Mediterranean climate: implications for tree decline diagnosis. Functional plant biology. Australian Journal Plant Physiologist, 28(3), 225-232. https://doi.org/10.1071/PP00119
García-Plazaola, J. I., & Becerril, J. M. (1999). A rapid HPLC method to measure lipophilic antioxidants in stressed plants: simultaneous determination of carotenoids and tocopherols. Phytochemical Analysis, 10(6),307-313
Jiménez, J. (1994). Los manglares del Pacífico Centroamericano. EFUNA.
Leipner, J., Frachebound, Y., & Stamp, P. (1999). Effect of growing season on the photosynthetic apparatus and leaf antioxidative defenses in two maize genotypes of different chilling tolerance. Environmental Experimental Botany, 42(2),129–139.
Lichtenthaler, H. K., Prenzel, U., Douce, R., & Joyard, J. (1981). Localization of prenylquinones in the envelope of spinach chloroplasts. Biochim. Biophys. 641(1), 99-105.
Lugo, A. E. (1981). The inland mangroves of Inagua. Journal of Natural History, 15(5), 845-852.
Ministerio del Ambiente y Recursos Naturales, Comanejo de Areas Protegidas, United States Agency for International Development y Fundación Nicaragüense para la Conservación y el Desarrollo. (2002). Plan de Manejo de la Reserva Natural Isla Juan Venado.
Munné-Bosch, S. (2005). The role of a-tocopherol in plant stress tolerance. Plant Physiology,162(7), 743–748. https://doi.org/10.1016/j.jplph.2005.04.022
Munné-Bosch S., & Alegre I. (2000). Changes in carotenoids, tocopherols and diterpenes during drought and recovery, and the biological significance of chlorophyll loss in Rosmarinus officinalis plants. Planta, (210), 925-931. https://doi.org/10.1007/s004250050699
Prahl, H. B. (1989). Manglares de Colombia. Villegas Editores.
Peng, C. L, Lin, Z. F., Su, Y. Z., Lin, G. Z., Dou, H. Y., & Zhao, C. X. (2006). The antioxidative funtions of lutein: electron spin resonance studies and chemical detection. Functional Plant Biology, 33(9), 839-846.
Sattler, S. E., Mene-Saffrane, L., Farmer, E. E., Kischke, M., Mueller, M. J., & DellaPenna, D. (2006) Nonenzymatic lipid peroxidation reprograms gene expression and activates defense markers in Arabidopsis tocopherol-deficient mutants. Plant Cell, 18(12), 3706–3720. https://doi.org/10.1105%2Ftpc.106.044065
Sussex, I. (1975). Growth and metabolism of the embryo and attached seedling of the viviparous mangrove, Rhizophora mangle L. American Journal of Botany, 62(9), 948-953. https://doi.org/10.2307/2441639
Stevens, W. D., Ulloa, C., Pool, A. y Montiel, O. M. (2001). Flora de Nicaragua. Introducción Gimnospermas y angiospermas. Missouri Botanical Garden.
Szymánska, R., & Kruk, J. (2008a). Tocopherol content and isomers’ composition in selected plant species. Plant Physiol Biochem, 46(1), 29–33. https://doi.org/10.1016/j.plaphy.2007.10.009
Szymánska, R., & Kruk, J. (2008b). Plastochromanol, a ‘new’ lipophilic antioxidant in Arabidopsis leaves, is synthesized by tocopherol cyclase. The effect of high-light on the level of prenyllipid antioxidants. En J. F. Allen, E. Gantt, J. H. Golbeck & B. Osmond (Eds.), Photosynthesis. Energy from the Sun: 14th International Congress on Photosynthesis (pp. 1579–1582). Springer
Tomlinson, P. B. (1999). The Botany of Mangroves. Cambridge University Press.
Yan, H., Zhang, P., Wang, C., Liu, Z., & Chang, W. (2007). Two lutein molecules in LHCII have different conformations and functions: Insights into the molecular mechanism of termal dissipation in plants. Biochemical and Biophysical Research Communications, 355(2), 457- 463. https://doi.org/10.1016/j.bbrc.2007.01.172
Notas de autor
1 Doctora
en Agrobiología Ambiental
2 Doctor
en Ciencias Biológicas
3 Doctora
en Vegetal y Ecología
Información adicional
Abreviaturas: A: Anteraxantina, Chl a: Clorofila
a, Chl b: Clorofila b, cm: Centímetro, DAP: Diámetro
a la altura del Pecho, E.S.: Error Estándar, Fv/Fm:
Máxima eficiencia fotoquímica del PSII , HPLC: (High Performance Liquid Chromatography), Cromatografía Liquida de Alta Resolución, L:
Luteína, Lx: Luteína epóxido, M: Molar, m: Metro, mm: Milímetro, N:
Neoxantina, nm: Nanómetro, ppm: Partes por millón, Toc: Tocoferoles, V: Violaxantina,
Z: Zeaxantina, VAZ: Violaxantina, Anteraxantina
y Zeaxantina, α-toc: Alfa-tocoferol, b-toc:
Beta-tocoferol, δ-toc: Delta-tocoferol, g-toc: Gamma-tocoferol,
µl: Microlitro.





