Abstract:
High stocking rate livestock management has determined the decline of native species of high forage value as Bromus auleticus Trin ex Nees in the Campos region of the Rio de la Plata grasslands. The species has some agronomic and ecological characteristics that define it as of high potential to be reintroduced in degraded natural grasslands. However, Bromus auleticus has not been sufficiently described, since commercial seeds and information regarding the establishment phase are scarce. Two field experiments were carried out in 2013 and 2014, reintroducing Bromus auleticus into native grasslands in the Basaltic region, aiming to evaluate the impact of sowing density in the forage productivity and their seasonal distribution. The treatments consisted of different sowing densities: 20, 40, 60 and 80 kg ha-1 and one control treatment. Considering the management and the evaluated period (4 years for experiments 2013, and 3 years for experiments 2014), different sowing densities did not modify the forage production in any of the plantings for the evaluated period and compared to the control. However, the use of sowing densities of 40 kg ha-1 or higher allowed modifying the seasonal distribution of forage production with a better contribution in autumn-winter, and more stable production throughout the year.
Keywords: Bromus auleticus, grassland, native species, planting density.
Resumen:
El manejo ganadero con altas cargas ha determinado la disminución de especies nativas de alto valor forrajero como el Bromus auleticus Trin. ex-Nees en la región Campos de los pastizales del Río de la Plata. La especie posee características agronómicas y ecológicas que la definen como de alto potencial para ser reintroducida en campos naturales degradados. Sin embargo, no ha sido lo suficientemente difundida por la falta de semilla comercial e información sobre la etapa de instalación. Se realizaron dos experimentos de campo en 2013 y 2014, a partir de la reintroducción de Bromus auleticus en cobertura sobre campo natural en la región de Basalto, con el objetivo de evaluar el efecto de la densidad de siembra en la producción de forraje y la distribución estacional. Los tratamientos consistieron en diferentes densidades de siembra: 20, 40, 60 y 80 kg ha-1 y un tratamiento testigo. Tomando en cuenta el manejo realizado y el período evaluado (4 años para el experimento 2013, y 3 años para el experimento 2014), la utilización de diferentes densidades de siembra no modificó la productividad total en ninguna de las siembras para el período evaluado y respecto al testigo. Sin embargo, la utilización de densidades de siembra iguales o superiores a 40 kg ha-1 modificó la distribución estacional de la producción de forraje hacia un mayor aporte en el período otoño-invernal y una producción más equilibrada durante el año.
Palabras clave: Bromus auleticus, campo natural, especies nativas, densidad de siembra.
Resumo:
O manejo da pecuária com altas cargas de animais tem determinado a diminuição de espécies nativas de alto valor forrageiro, como Bromus auleticus Trin. ex Nees na região do Bioma Campos. A espécie possui características agronômicas e ecológicas que a definem como tendo alto potencial para ser reintroduzida em campos naturais degradados. No entanto, não foi suficientemente estudado devido à falta de sementes comerciais e informações sobre a fase de instalação. Dois experimentos de campo foram realizados em 2013 e em 2014 a partir da reintrodução de Bromus auleticus em cobertura em campo natural na região do Basalto, com o objetivo de avaliar o efeito da densidade de semeadura na produção de forragem e sua distribuição sazonal. Os tratamentos consistiram em diferentes densidades de semeadura: 20, 40, 60 e 80 kg ha-1 e um tratamento testemunha sem plantio. Considerando o manejo realizado e o período avaliado (4 anos para o experimento de 2013 e 3 anos para o experimento de 2014), o uso de diferentes densidades de semeadura não alterou a produtividade total de forragem em nenhum dos experimentos para o período avaliado e com relação ao controle. Porém, a utilização de densidades de semeadura iguais ou superiores a 40 kg ha-1 permitiu modificar a distribuição sazonal da produção forrageira com maior contribuição no período outono-inverno e produção mais equilibrada ao longo do ano.
Palavras-chave: Bromus auleticus, campo nativo, espécies nativas, densidade de semeadura.
Carátula del artículo
Effect of sowing density on the productivity of a native grassland restored with Bromus auleticus
Efecto de la densidad de siembra en la productividad
de un campo natural restaurado con Bromus auleticus
Efeito da densidade de semeadura na productividade
de um campo nativo restaurado com Bromus auleticus
 Franco
Franco,
R. rreyno@inia.org.uy
Universidad de la República, Uruguay
Jaurena,
M.
Instituto Nacional de Investigación Agropecuaria
(INIA), Uruguay
Reyno,
R.
Instituto Nacional de Investigación Agropecuaria
(INIA), Uruguay
Agrociencia Uruguay
Universidad de la República, Uruguay
ISSN-e: 2730-5066
Periodicity: Bianual
vol. 25, no. 2, e333, 2021
Received: 25 February 2020
Accepted: 01 July 2021
Published: 06 September 2021
Corresponding author: rreyno@inia.org.uy
1. Introduction
In the Campos biome region, overgrazing is one of the main causes of ecosystem degradation(1). Natural fields are repeatedly exposed to overgrazing, especially when coinciding with periods of low forage growth (for example winters or severe droughts). In both natural and cultivated pastures, seasonal variations in forage production are related to fluctuations in temperature, humidity, radiation, and grazing pressure(2)(3)(4). In the autumn-winter period, forage production is minimal due to low temperatures and frosts that induce the physiological dormancy of the dominant native C4 grasses in most natural grasslands(4)(5). Therefore, overgrazing under these conditions can lead to a reduction in pasture growth(6) and species diversity(7). In this context, the reintroduction of winter perennials together with the addition of nutrients and the management of the pasture with strategic breaks and control of the forage supply would allow the productive recovery of fields degraded by overgrazing, especially in the autumn-winter period(8)(9)(10)(11).
The traditional "improvement" technology of the natural grassland, that is, the introduction of species in the natural grassland, is a strategy that seeks to increase productivity without affecting the integrity of the ecosystem(12)(13). In the Campos biome, the main improvement strategies are based on planting exotic coverage species, mainly legumes of the Lotus and Trifolium genera, together with nitrogen and/or phosphate fertilization. To favor the establishment of the species to be introduced, in addition to reducing the competition of the resident vegetation, it is necessary to have dry remains and mulch, which protect the seeds by preserving humidity and avoiding drastic temperature fluctuations at ground level(12)(14)(15). In the first years after the improvements, primary and secondary productivity increase, but in the medium term, some sown species do not persist, leaving spaces that are generally occupied by other exotic species(16).
Bromus auleticus Trinius is a winter perennial species of the Festuceae tribe native to southern Brazil, Uruguay, and Argentina(17)(18)(19), and one of the most promising species for the recovery of degraded natural grasslands. This species has the following ecological advantages for its introduction or reintroduction: i) adaptation to cover crops(14)(20)(21); ii) autumn-winter contribution of high nutritional value(22)(23)(24); iii) adaptation to situations of low fertility and tolerance to summer water stress(2)(25), iv) spontaneous reseeding capacity and productive stability(2)(9)(23). The species also has a great diversity of ecotypes adapted to different edaphoclimatic regions, which is an opportunity to select local ecotypes(26).
Despite all its favorable characteristics, the slow implantation of Bromus auleticus is a limiting factor for the initial production of the species, affecting the conditions for competition, its survival, and future persistence(17)(22)(27). In this regard, the improvement in the initial establishment could reduce the time without grazing, which would facilitate its use for the recovery of degraded fields. Experiences of planting native grass covers on surface soils aimed at the productive and ecological recovery of natural grasslands have been scarce in the region. Sowing density is among the most important factors that affect the establishment of the species(3)(10). The sowing densities used for Bromus auleticus in previous research range from 10 to 60 kg ha-1, both in monoculture(2)(17)(28) and in natural grassland coverage(9)(11)(14). In cover plantings, forage yield increases due to the initial increase in plants and tillers achieved by using greater sowing densities are reduced with time(29), as was observed in conventional Bromus plantings(28). According to Carámbula(3), the plant population variation has a compensatory effect in later stages and under cultivation conditions, maintaining the yield due to the variation in their weight and size. The maximum levels of forage production of the species are only reached after the second or third year of life(24)(30)(31). In any case, the relationship between the number of implanted plants and the initial productivity is not clear. Consequently, it is necessary to study the factors that affect the initial productivity of Bromus auleticus in cover plantings on natural grasslands.
In this way, it is hypothesized that increasing the sowing density of Bromus auleticus cover increases the number of plants, which allows increasing winter and total production in natural grassland improvements. Therefore, this study aims to evaluate the effect of different sowing densities of Bromus auleticus on the seasonality and forage production of the natural grassland.
2. Materials and methods
2.1 Plant material and preparation of the experimental
area
The
research was carried out at the Glencoe Experimental Unit (INIA) in northern
Uruguay (32.09´S; 57.81´W). Two field experiments were carried out, Experiment
2013 (E2013) and Experiment 2014 (E2014), in which the effect of the sowing
density of Bromus auleticus cover on a natural
grassland with Basalt soil was evaluated. An experimental line of Bromus auleticus was used, generated by the INIA Pastures and
Forages Program, selected from a Basalt ecotype experimentally called “Los Paraísos”. The sown seed corresponded to the 2012 and 2013
harvests for E2013 and E2014, respectively. The first experiment was
established on May 7, 2013 (E2013), and the second on May 28, 2014 (E2014). The
experimental sites were located in adjacent areas; however, differences were
detected in the average soil depth: 31 cm and over 50 cm for E2013 and E2014,
respectively. The 10 main species with the highest coverage in the site, where
both experiments were carried out, were Aristida uruguayensis Henrard., Baccharis coridifolia DC., Bothriochloa laguroides (DC) Herther, Coelorhachis selloana (Hack.de Koning & Sosef), Eragrostis lugens Nees, Nasella nessiana (Trin. & Rupr.) Barkworth, Paspalum
notatum Flüggé, Piptochaetium
stipoides (Trin. & Rupr.)
Hack. ex Arechav, Schizachyirium spicatum (Spreng.) Herter and some sedges. While
the soil cover was: dry remains 40%, green fraction 56%, bare soil 2%, and
stone 2%. The experimental area was prepared by cutting with an experimental
grass cutter at a height of 3 cm. Before the cuttings, the sites were not
grazed for about 120 days. This management sought to increase the fractions of
dry remains and mulch in the experimental area.
2.2
Climatic conditions
Climate
data were obtained based on the Glencoe INIA Weather Station. The Percentage of
Available Water in the Soil (PAW) was estimated with the climatic data using 60
mm and 100 mm sheets for E2013 and E2014, respectively, according to the type
and depth characteristics of the soil. The PAW measures the percentage of water
available in the profile, in relation to the field capacity (100%). The
available water content estimation was carried out with a spreadsheet prepared
by the INIA GRAS Unit that considers the type of soil, the effective rainfall,
potential atmospheric water demand and plant transpiration(32).
2.3 Experimental design and treatments
A
randomized complete block design with three replications was used. The
experimental units were 2×5 m plots broadcast on a Basalt natural grassland.
The treatments consisted of four sowing densities: 20 (T1), 40 (T2), 60 (T3)
and 80 (T4) kg ha-1 of Bromus auleticus
seeds (with germination percentages of 74 and 76% for E2013 and E2014,
respectively), and a non-seeded control treatment, fertilized (FNG, fertilized
natural grassland) with the same dose as the seeded treatments. Before the
sowing, fertilization was carried out with 60 kg ha-1 of P205
and 100 days after sowing (DAS) it consisted of 30 kg ha-1 of N (as
urea). In the following years, the treatments were refertilized
with 30 kg ha-1 of N (as urea) in autumn, in both experiments. The
used seeds were previously treated with TMTD fungicide + Carbendazim (0.02 ml
kg-1 of seed) and Tiamethoxan insecticide
(0.02 ml kg-1 of seed).
2.4 Evaluated variables
The number of plants per m-2 at 365 DAS was estimated by direct counting using 3 fixed rectangles of 0.1 m2 in each experimental unit. Subsequently, the total accumulated forage production (DM kg ha-1) corresponding to the sum of the production of the second and third years in each experiment (5 and 8 cuttings in experiments 2013 and 2014, respectively) and the seasonal forage production (DM kg ha-1) were estimated. Forage production was determined using a Honda cutter with a collection bag (model HRC 216, Japan), at a height of 10 cm, in a 0.52 m strip (in the center of each experimental unit) for 5 m long, which represented a sampling area of 2.6 m2. Samples were dried in a forced-air oven at 60 ℃ for 72 hours, and the dry matter content was determined by weighing on a precision scale.
There were no cuttings or grazing in either of the experiments from sowing until the final cuttings of the first summer (remnant of 10 cm) carried out on 2/25/2014 and 2/25/2015, respectively. These were not included in the determination of accumulated or seasonal forage production. The forage cuttings made to estimate the total accumulated forage production are presented in Table 1.
Table 1
Experiment, cutting date, and growth
days for the estimation of accumulated forage production
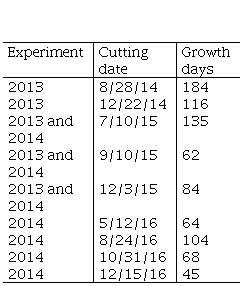
The seasonal forage production was estimated (DM kg ha-1) from the 2016 cuttings of both experiments, corresponding to the fourth and third years of the pastures (E2013 and E2014, respectively). Additionally, the percentage contribution (%) of seasonal forage was calculated for each experiment according to the total annual forage corresponding to 2016.
Finally, the plant coverage of Bromus auleticus was determined in winter 2014 only for E2013, by visual estimation (%), through 5 observations (subsamples) using a 50×50 cm section, in a transect for each experimental unit, equidistant between observations. This variable was not estimated for E2014.
2.5 Statistical analysis
The number of plants per m-2 a year after
sowing each experiment was estimated using a mixed generalized linear model,
which was fitted with a Poisson distribution with log link function. The
fixed effects included in the model corresponded to block, density and
experiment effects. The associations with the contribution (%) of seasonal
forage were then made with these estimates. The accumulated forage production
of the natural grassland (DM kg ha-1) was analyzed using a mixed
general linear model, which was adjusted to a normal distribution. The effects
included in the model correspond to the fixed effects: block, treatment,
experiment, and treatment × experiment interaction. Based on this and the
presence of the "experiment" effect (and no experiment × treatment
interaction), a mixed general linear model was used adjusting a normal
distribution and including block and treatments as fixed effects for the
analysis of seasonal forage production (DM kg ha-1), and the
percentage contribution of seasonal forage (%) for each experiment. For significant
effects, the adjusted means of the treatments were compared with Fisher's LSD
measurement comparison test (α = 0.05). To estimate the relationship between
the number of plants per m-2 a year after sowing and the
contribution (%) of seasonal forage, a simple linear regression analysis (y=a+bx) was used, estimating the coefficients of the function
and the determination coefficient (R2), in each case. To analyze the
effect of the treatments on the proportion of Bromus auleticus,
a mixed general linear model was used, fitting a normal distribution. The
effects included in the model correspond to the fixed effects: block and
treatments, and each subsample as random effects. The InfoStat(33)
software was used for the analysis of the variables, with its interface with
the R software(34).
3. Results and discussion
3.1 Climatic conditions
Figure
1 shows the evolution of the percentage of available water in the soil (PAW)
and the daily rainfall that occurred throughout the experiment (partitioned by
year) in both experiments.
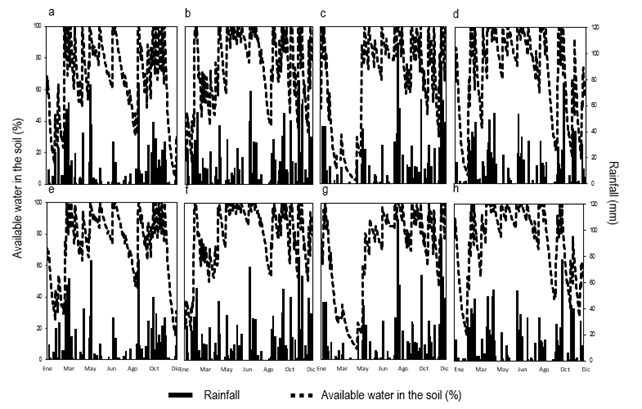 Figure 1
Estimation of available water in the
soil (%) (Main axis) and rainfall record (mm) (Secondary axis) according to
month of the year: a, b, c and d corresponding to 2013, 2014, 2015 and 2016, at
the Experiment site 2013, and e, f, g and h corresponding to 2013, 2014, 2015
and 2016, at the Experiment site 2014, respectively. Own elaboration based on
data provided by INIA Gras based on the Glencoe Meteorological Station
Figure 1
Estimation of available water in the
soil (%) (Main axis) and rainfall record (mm) (Secondary axis) according to
month of the year: a, b, c and d corresponding to 2013, 2014, 2015 and 2016, at
the Experiment site 2013, and e, f, g and h corresponding to 2013, 2014, 2015
and 2016, at the Experiment site 2014, respectively. Own elaboration based on
data provided by INIA Gras based on the Glencoe Meteorological Station
The PAW remained equal to or greater
than 60%, in the 120 days after the establishment of E2013, except in August,
which was around 30%. For this experiment, the first summer presented a relatively short period with
low levels of PAW. In E2014, the PAW remained above 80% in most of the period
between sowing and 120 days after sowing, however, and unlike in E2013, the PAW
in the first summer was lower than 30%, specifically in February and March. It
should be noted that this long period (February and March 2015) coincides with
the second summer for E2013 and the first summer for E2014. For the stage
following the establishment of both experiments, there were periods of autumn,
winter, and spring with PAW above 60 and 80% for E2013 and E2014, respectively,
except for the previously mentioned 2015 period. On the other hand, the high
rainfall record in autumn 2016 stands out, generating high PAW in both
experiences, as well as the record of 13 meteorological frost events in June
2016 higher than the historical record.
3.2 Accumulated forage production
In the accumulated forage production, which totals 581 days of growth for E2013 and 562 days for E2014, a strong effect of the experiment was observed (p<0.0001), without interaction between treatments and experiment (p=0.88) (Table 2). When performing the analysis separately, significant differences were observed between treatments only in E2014 (p=0.04).
Accumulated forage production (2nd + 3rd pasture year) in E2014 was higher than in E2013, which would be related to differences in the average depth of soils (31 and over 50 cm for sites 2013 and 2014, respectively) and to the production of natural grassland species as they are different experimental sites. Additionally, in the 3rd year of E2014 (corresponding to 2016, which does not affect the accumulated of E2013), a period with high levels of PAW occurred between autumn and winter, as a result of a high rainfall record (725 mm) despite having had 13 meteorological frost events in June of the same year. In E2014, treatments T2 and T3 had higher accumulated forage production, however, they did not differ from FNG. On the other hand, the inferiority of both T1 and T4 could be due to the fact that they were still below (T1) or above (T4) the maximum response capacity of the environment, in a context of abiotic (such as water stress) and biotic interactions of intra and interspecific competition(35)(36).
Table 2
Accumulated forage production (DM kg
ha-1), according to density and experiment corresponding to the 2nd
+ 3rd year, for each experiment
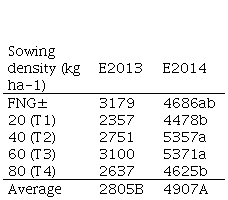 ± Fertilized natural grassland
± Fertilized natural grassland Different uppercase and lowercase letters mean significant differences (p <0.05).
A
14% contribution of the species was recorded in cover plantings with partial
tapestry removal, on Cristalino soils, in the spring
of the year after sowing(37).
The following year (3rd year of improvement)(38)
presented an increase in the contribution of Bromus auleticus (40%), in the same field improvement. On the other hand, Carámbula and others(39)
observed on Cristalino subeutric argisols an accumulated production (3 years) of the
Natural Field of 20448 DM kg ha-1 with an accumulated contribution
of Bromus auleticus of 463 DM kg ha (2.2%).
3.3 Seasonal forage production
Bromus auleticus coverage in winter 2014 (only for E2013) was of 7, 11, 14 and 16 (%), T1, T2, T3 and T4, respectively, with lower Bromus coverage when densities were lower (p=0.008) compared to higher densities (T3 and T4).
Regarding the production of seasonal forage (DM kg ha-1), there were differences between treatments only in winter in E2013 (p=0.02), while in E2014 the differences between treatments in summer (p=0.01) and winter (p=0.05) stand out, with no significant differences in autumn and spring (p=0.06 in both seasons) (Figure 2). In winter, the forage production of the sown treatments increased 64% (E2013) and 39% (E2014) on average compared to the control (Figure 2).
Additionally, in both experiments, forage production in winter increased significantly from 40 kg ha-1 on, above which there are no additional increases.
Significant differences (p<0.01) were generated in the summer in favor of the control, in E2014. In this situation, the sown treatments had an average forage yield 28% lower than that of the control, and 37% lower in the sown treatment of Bromus with 80 kg ha-1. The winter forage production in T2 was 144 and 221 DM kg ha-1 for E2013 and E2014, respectively, while the control forage production for the same season was 82 and 157 DM kg ha-1, respectively, which shows the superiority in forage production due to the inclusion of the species in this period (p=0.016 and p=0.01, for 2013 and 2014, respectively).
These seasonal production results are lower compared to previous studies by García(2) in conventional Bromus auleticus plantings using different genetic material and in grasslands planted in high potential soils. However, in the case of improvements with Bromus auleticus, Castrillón and Pirez(40) estimated availability of 3300 DM kg ha-1 in March, corresponding to the first summer of the pasture and a corresponding availability to the autumn-winter growth of 3760 DM kg ha-1 of the second year of the pasture with a contribution of Bromus auleticus of 11%, similar to this experience.
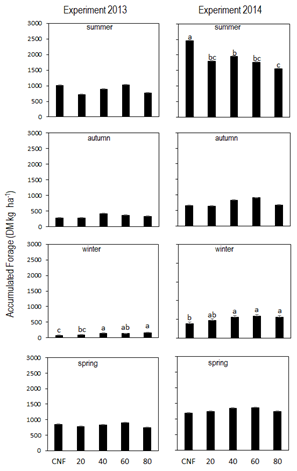 Figure 2
Seasonal forage production (DM kg ha-1)
according to treatments: a, b, c and d corresponding to summer, autumn, winter
and spring for E2013, and e, f, g and h corresponding to summer, autumn, winter
and spring for E2014, respectively
Figure 2
Seasonal forage production (DM kg ha-1)
according to treatments: a, b, c and d corresponding to summer, autumn, winter
and spring for E2013, and e, f, g and h corresponding to summer, autumn, winter
and spring for E2014, respectively
Different lowercase letters in the same station and experiment
correspond to statistically significant differences (p<0.05).
Regarding
the seasonal distribution of annual forage production, significant differences
were detected for all seasons in E2014 (p<0.05) (Figure 3), while no
differences were observed in E2013. The inclusion of the species generated
increases in the autumn-winter proportion until T2 and a reduction
of the summer contribution regarding the FNG (p<0.05).
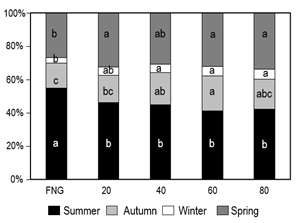 Figure 3
Seasonal distribution of percentage
forage production (%) according to treatments for E2014
Figure 3
Seasonal distribution of percentage
forage production (%) according to treatments for E2014
Different lowercase letters correspond to significant
differences between treatments for the same season (p<0.05).
In E2014, the FNG presented a forage distribution (%) of 55(V), 15(0), 3(I) and 27(P), while in T2 (not statistically different at higher densities) the distribution (%) was 45, 19, 5 and 31 for the same stations. For Basalt natural grasslands, fertilized with nitrogen and phosphorus distributed between the beginning of autumn and the end of winter, average distributions (%) of 50(V), 16(O), 11(I) and 23(P)(41) and 35(V), 27(O), 11(I) and 27(P)(4) were obtained. This differs to the findings of this experiment, although they are similar in terms of intra-annual variation with a concentration in the spring-summer period. The seasonal distribution (%) of a field improvement with Trifolium repens and Lotus corniculatus observed by Ayala and others(42) in the third year on Cristalino soils was 28(V), 18(O), 13(I) and 41(P), similar to the distribution observed with these same species in the Glencoe experimental station (site of this experience) in an average of 6 years: 24(V), 19(0), 17(I) and 40(P)(43).
The interaction of different factors such as the forage collection height used in this study (10 cm), the prostrate growth habit of the ecotype planted, as well as unfavorable climatic conditions for growth in winter 2016 probably underestimated productivity, mainly in winter. Likewise, the use of high remnants would allow the species to increase the production of aboveground dry matter and roots, impacting its persistence(44)(45), and contemplate the low tolerance to high levels of defoliation(46).
The lower contribution in the summer of the treatments sown compared to the FNG, reflected both in terms of seasonal production (DM kg ha-1) and distribution of annual forage production (%), could be explained by an increase in the ratio of C3/C4 species of the natural grassland in the sown plots and compared to the treatments without plantings, with a possible interference of the introduced grass on the potential growth of the dominant summer native grasses (C4) of the natural grassland.
In summary, the higher contribution of Bromus auleticus in autumn-winter and the lower relative contribution in the summer are modifying the typical forage production curve of the natural grassland of Basalt soils, environments that are characterized by high spring-summer growth peaks and low autumn-winter contributions(4). The differences in the seasonal distribution could be associated with a greater presence of Bromus auleticus in environments with greater productive potential, as occurred in E2014.
It should be noted that these results come from the third and fourth year after sowing in 2014 and 2013, respectively, indicating the permanence of the effect of sowing density on forage production, similar to that found in some studies on conventional sowing(47) and coinciding with what Moliterno and others(28) highlighted in terms of the increase observed in productive stability over time in Bromus auleticus plantings.
3.4 Relationship between the number of plants per m-2
a year after sowing and the contribution to seasonal forage production
In
E2013, significant associations were found between the number of plants per m-2
of Bromus auleticus observed a year after
sowing and the contribution to the forage production in summer (p=0.036),
autumn (p=0.001) and winter (p=0.002), while there were no significant
relationships in spring (p>0.05) (Figure 4, Graphs b, c, a and d, respectively).
In the case of E2014, there were associations with the seasonal contribution
(%) in all seasons: summer (p=0.0001), autumn (p=0.002), winter (p=0.012) and
spring (p=0.03) (Figure 4, Graphs e, f, g and h, respectively).
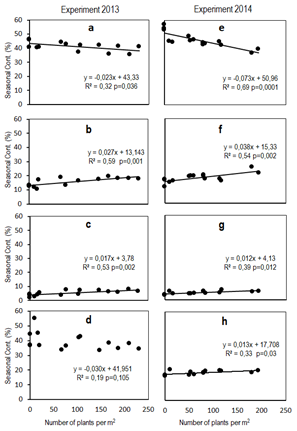 Figure 4
Relationship between the number of
plants of Bromus auleticus per m-2
a year after sowing with the seasonality of forage production: a, b, c and d
correspond to summer, autumn, winter and spring of E2013, and e, f, g and h
correspond to summer, autumn, winter and spring of E2014, respectively
Figure 4
Relationship between the number of
plants of Bromus auleticus per m-2
a year after sowing with the seasonality of forage production: a, b, c and d
correspond to summer, autumn, winter and spring of E2013, and e, f, g and h
correspond to summer, autumn, winter and spring of E2014, respectively
In general terms, the contribution in autumn increased from 12% to 25%, while in winter, from 3 to 8% as the number of plants per m2 a year after sowing increased from 0 plants to more than 150 plants per m2.
On the other hand, both experiments presented a negative relationship between the increase in the number of plants of Bromus auleticus and the proportion of summer-produced forage, which would be related to the lower growth of the summer species as stated in the previous point. Therefore, it is noteworthy that the increase in the number of Bromus auleticus plants in E2014 would explain almost 69% of the changes in the seasonal contribution of forage production in summer. This effect could be related to the competence exercised by Bromus auleticus with its caespitose growth habit on the production of dominant summer perennial grasses. In addition to the positive impact on autumn-winter productivity, the increase in the density of Bromus auleticus could also improve the nutritional quality due to the high nutritional value of the species(22)(23)(24).
It is important to note that effects were observed in the 2nd and 3rd year after sowing, which shows that the changes were persistent. Davies(23) points out the advantage of planting at high densities to increase the competition exerted by light. But this effect would be lost after the second year, possibly related to plasticity mechanisms that lead to size-density compensations reported by Lemaire and Chapman(48), and/or due to higher Bromus auleticus plant mortality in the high-density treatments after the first year. High plant density possibly increased the biotic interactions of intraspecific and interspecific competition, and, therefore, higher plant mortality in the context of abiotic restrictions, as in the case of water stress(35)(36). However, the persistence of the effects of the Bromus auleticus sowing density over time implies that the size-density compensations were very scarce or null.
Formoso(49) and Zarza and others(50) found high associations between sowing density, the number of plants and the forage production of the first year when working with Festuca (winter perennial grass) in conventional plantings. However, due to the greater flexibility of perennials to different factors in the long term (3-4 years) and the tillering process, this association decreases with time. In the present experiment, the increase in the sowing density of Bromus auleticus persistently improved the autumn-winter forage production, seasons in which the natural grasslands of the region present minimum productivity values. Given the impact that the improvement in production and winter quality of forage has in livestock systems based on natural grasslands(51), it is necessary to deepen the study of the effect of the sowing density in new experiments.
4. Conclusions
The inclusion of Bromus auleticus cover, regardless of the sowing density, did not modify the total forage production compared to the control treatment. However, the increase in sowing density up to 40 kg ha-1 allowed modifying the seasonality of forage production in favor of the autumn-winter period in both experiments, and also a lower forage production in summer in the site with the greatest potential.
The number of Bromus auleticus plants a year after sowing was directly related to an increase in autumn-winter forage production, an effect that persisted through the 3rd and 4th years of the pasture.
Additional information
Authors' contribution: Rodolfo Franco contributed with
experimental design, data collection, interpretation and analysis of the
information, and writing of the article. Martín Jaurena
and Rafael Reyno contributed with experimental
design, interpretation and analysis of the information, and writing of the
article.
Editor: The
following editor approved this article.
Sebastián Mazzilli (https://orcid.org/0000-0001-7883-7124)
Universidad
de la República Paysandú, Uruguay.
Acknowledgments
To
the officials and workers of the Glencoe Experimental Station (INIA Tacuarembó)
for their collaboration in conducting the experiments.
References
1. Tiscornia G, Jaurena M, Baethgen W. Drivers, process, and consequences of native grassland degradation: Insights from a literature review and a survey in Río de la Plata grasslands. Agronomy [Internet]. 2019 [cited 2021 Jul 07];9(5):239. Available from: https://bit.ly/3hpoaTm.
2. García J. Crecimiento y calidad de gramíneas forrajeras en la Estanzuela. Montevideo: INIA; 2003. 35p.
3. Carámbula M. Pasturas y Forrajes: insumos, implantación y manejo de pasturas. Montevideo: Hemisferio Sur; 2003. 371p.
4. Berretta EJ. Principales características de las vegetaciones de los campos de Basalto. In: Berretta EJ, editor. Reunión del grupo técnico regional del cono sur en mejoramiento y utilización de los recursos forrajeros del área tropical y subtropical: grupo campos. Montevideo: INIA; 1998. p. 11-9.
5. Oliveira JCP, Moraes COC. Distribuição da produção equalidade de forragem de Bromus auleticus Trinius. Pesqui Agropecu Bras. 1993;28(3):391-8.
6. Pallarés OR, Berretta EJ, Maraschin GE. The South American campos ecosystem. In: Suttie J, Reynolds SG, Batello C, editors. Grasslands of the World. Roma: FAO; 2005. p. 171-219.
7. Fedrigo JK, Ataide PF, Filho JA, Oliveira LV, Jaurena M, Laca EA, Overbeck GE, Nabinger C. Temporary grazing exclusion promotes rapid recovery of species richness and productivity in a long‐term overgrazed Campos grassland. Restor Ecol. 2018;26(4):677-85.
8. Jaurena M, Bentancur O, Ayala W, Rivas M. Especies indicadoras y estructura de praderas naturales de basalto con cargas contrastantes de ovinos. Agrociencia Uruguay. 2011;15(1):103-14.
9. Boggiano P, Zanoniani R, Saldanha S. Implantación de Bromus auleticus Trin. en cobertura. In: Reunião temática internacional sobre o gênero Bromus. Bagé (RS): Embrapa Pecuária Sul; 2000. p. 12-3.
10. Silva GM, Maia MS, Moraes COC. Efeito da população de plantas sobre o rendimento e a qualidade de sementes de cevadilha-vacariana (Bromus auleticus Trinius). In: Los recursos fitogenéticos del género Bromus en el Cono Sur. Montevideo: PROCISUR; 2001. p. 77-80.
11. Ayala W, Bermúdez R, Ferrés S. Evaluación de diferentes gramíneas para mejoramientos de campo sobre suelos de sierra. In: Jornada Pasturas en la sierra: Sitio Experimental Arbolito. Treinta y Tres: INIA; 2003. p. 4-10.
12. Risso D. Siembras en el tapiz: consideraciones generales y estado actual de la información en la zona de suelos sobre Cristalino. In: Carámbula M, Vaz Martins D, Indarte E, editors. Pasturas y producción animal en áreas de ganadería extensiva. Montevideo: INIA; 1997. p. 71-82.
13. Jacques AVÁ, Heringer I, Scheffer-basso SM. Aspectos do manejo e melhoramiento da pastagem nativa. In: Pillar VP, Müller SC, Castilhos ZMS, Jacques AVA, editors. Campos Sulinos: conservacao e uso sustentavel da biodiversidade. Brasilia: MMA; 2009. p. 237-47.
14. Ayala W, Carámbula M. Gramíneas para mejoramientos extensivos. In: Mas C, Carámbula M, Bermúdez R, Ayala W, Carriquiry E, editors. Mejoramientos extensivos en la región este: resultados experimentales 1991-92. Treinta y Tres: INIA; 1992. p. 39-48.
15. Dowling PM, Clements RJ, McWiliam JR. Establishment and survival of pasture species from seeds sown on the soil surface. Aust J Agric Res. 1971; 22(1):61-74.
16. Jaurena M, Lezama F, Salvo L, Cardozo G, Ayala W, Terra J, Nabinger C. The dilema of improving native grasslands by overseeding legumes: production intensification or diversity conservation. Rangel Ecol Manag. 2016;69(1):35-42.
17. Olmos F. Bromus auleticus. Montevideo: INIA; 1993. 30p.
18. Moraes COC, Oliveira JCP, Paim NR. Comparação de Bromus auleticus Trinius com outras gramíneas perenes de inverno. Bagé (RS): Embrapa Pecuária Sul; 2000. 30p.
19. Millot JC. Bromus auleticus: una nueva especie domesticada. In: Los recursos fitogenéticos del género Bromus en el Cono Sur. Montevideo: PROCISUR; 2001. p. 3-6.
20. Carriquiry E, Majó G, Saldanha S, Millot JC. Bromus auleticus: efecto de la fertilización y manejo del pastoreo sobre producción de semillas y sus componentes. In: II Seminario de Campo Natural. Montevideo: Hemisferio Sur; 1990. p. 89-104.
21. Zanoniani R, Boggiano P, Saldanha S. Implantación de Bromus auleticus Trinius en cobertura. In: Los recursos fitogenéticos del género Bromus en el Cono Sur. Montevideo: PROCISUR; 2001. p. 35-8.
22. Moraes COC, Oliveira JCP, Paim NR. Comparação de Bromus auleticus Trinius com outras gramíneas perenes de inverno. Bagé (RS): Embrapa Pecuária Sul; 2000. 30p.
23. Davies P. Efecto del nivel de nitrógeno y densidad de siembra en Bromus auleticus. In: II Seminario Nacional de Campo Natural. Tacuarembó: Hemisferio Sur; 1990. p. 105-14.
24. Bemhaja M. Gramínea nativa perenne invernal para suelos arenosos: Bromus auleticus cv INIA Tabobá. In: Los recursos fitogenéticos del género Bromus en el Cono Sur. Montevideo: PROCISUR; 2001. p. 103-4.
25. Formoso FA, Allegri MA. Gramíneas Perennes en el noreste. Montevideo: CIIAB; 1984. 37p.
26. Condón F, Jaurena M, Reyno R, Otaño C, Lattanzi F. Spatial analysis of genetic diversity in a comprehensive collection of the native grass Bromus auleticus Trinius (ex Nees) in Uruguay. Grass Forage Sci. 2017;72(4):1-11.
27. Silva GM, Moraes COC. Cevadilha Vacariana (Bromus auleticus Trinius): histórico, utilização e perspectivas. Bagé (RS): Embrapa Pecuária Sul; 2009. 20p.
28. Moliterno E, Saldanha S, Rucks F. Establecimiento y producción inicial de mezclas de dos cultivares de Bromus auleticus y uno de Festuca arundinacia con leguminosas. In: Los recursos fitogenéticos del género Bromus en el Cono Sur. Montevideo: PROCISUR; 2001. p. 87-95.
29. Carámbula M. Pasturas naturales mejoradas. Montevideo: Hemisferio Sur; 1996. 524p.
30. Rosengurtt B. Gramíneas y leguminosas de Juan Jackson. Comportamiento en el campo y en ensayos de cultivo. In: Rosengurtt B, editor. Estudios sobre praderas naturales del Uruguay: 5a contribución. Montevideo: Rosgal; 1946. p. 215-346.
31. Boggiano P. Evaluación de 14 gramíneas perennes bajo pastoreo. In: II Seminario Nacional de Campo Natural. Tacuarembó: Hemisferio Sur; 1990. p. 185-95.
32. INIA. Modelo logarítmico BH [Internet]. Montevideo: INIA; c2021 [cited 2021 Jul 07]. Available from: https://bit.ly/3bJdImJ.
33. Di Rienzo JA, Casanoves F, Balzarini M, Gonzalez L, Tablada M, Robledo CW. InfoStat [Internet]. Versión 2017. Córdoba: Grupo Infostat; 2017 [cited 2021 Jul 07]. Available from: https://bit.ly/3dDvIyu.
34. R Development Core Team. R: a language and environment for statistical computing [Internet]. Vienna: R Foundation for Statistical Computing; 2016 [cited 2021 Jul 07]. Available from: https://bit.ly/31v5U2G.
35. Harper J. Population biology of plants. New York: Academic Press; 1977. 892p.
36. Cook SJ. Establishing pasture species in existing swards: a review. Trop Grassl. 1980;14:181-7.
37. Donagaray F, Lecueder F. Siembra de pastos nativos en cobertura de 1981, analizada durante 1982-83 [grade’s thesis]. Montevideo (UY): Universidad de la República, Facultad de Agronomía; 1984. 89p.
38. Inciarte JC, Rienzi M. Siembra de pastos nativos en cobertura de 1981 y comparación con campo natural pastoreado, analizada durante 1983-84 [grade’s thesis]. Montevideo (UY): Universidad de la República, Facultad de Agronomía; 1984. 136p.
39. Carámbula M, Bermúdez R, Ayala W. Evaluación de gramíneas promisorias para mejoramientos extensivos. In: Jornada anual de producción animal: resultados experimentales 1997-1998. Treinta y Tres: INIA; 1998. p.1-11.
40. Castrillón A, Pirez C. Evaluación de la capacidad de instalarse de especies forrajeras en el campo natural con diferentes tratamientos de laboreo [grade’s thesis]. Montevideo (UY): Universidad de la República, Facultad de Agronomía; 1987. 264p.
41. Berretta EJ, Risso D, Levratto J, Zamit W, Zarza M. Evolución y producción de vegetaciones naturales de Basalto con fertilización NP. In: Tecnologías de producción ganadera para Basalto. Tacuarembó: INIA; 1997. p. 7-11.
42. Ayala W, Bermúdez R, Machado C, Nuñez I, Gonzales S, Monteagudo M. Aspectos relevantes para mejorar la productividad y persistencia de pasturas mejoradas: el caso de mejoramientos de campo en base a Trébol Blanco-Lotus. In: Jornada Anual de Producción animal: resultados experimentales. Treinta y Tres: INIA; 2002. p. 7-31.
43. INIA. Visita grupos CREA. Tacuarembó: INIA; 1996. 24p.
44. Scheffer-basso SM, Soares GC, Dall’agnol M. Efeito de freqüência e altura de corte em dois genótipos de Bromus auleticus trin, ex ness. Rev bras agrocienc. 2002;8(3):191-4.
45. Santos DS, Branco VTA, Mazzocato AC, Ferreira JL. Desempenho de três acessos de Bromus auleticus com relação ao efeito de alturas de cortes [Internet]. 2012 [cited 2021 Jul 07]. Available from: https://bit.ly/3jT2TU0.
46. Zanoniani R, Boggiano P, Cadenazzi M. Respuesta poblacional invernal de dos gramíneas nativas a la fertilización nitrogenada y oferta de forraje. Agrociencia Uruguay. 2012;16(2):103-9.
47. Silva RDD, Scuro VM, Silva EF da, Lima LC de, Oliveira JCP. Efeito da densidade de semeadura na produção de matéria seca de Bromus auleticus Trinius. In: 15ª Mostra da Produção Universitaria. Rio Grande (BR): UFR; 2016. p. 2.
48. Lemaire G, Chapman D. Tissue flows in grazed plants communities. In: Hodgson J, Illius AW, editors. The ecology and management of grazing systems. Wallingford: CAB International; 1996. p. 3-37.
49. Formoso F. Festuca arundinacea, manejo para producción de forraje y semillas [internet]. Montevideo: INIA; 2010 [cited 2021 Jul 07]. 192p. Available from: https://bit.ly/3xodqul.
50. Zarza R, Calistro E, Martinez E. Importancia de la implantación en las pasturas perennes. In: Reunión técnica: el éxito productivo de una pastura con leguminosas perennes comienza en su implantación. Colonia: INIA; 2013. p. 1-9.
51. Jaurena M, Durante M, Devincenzi T, Savian V, Bendersky D, Moojen FG, Pereira M, Soca P, Quadros FL, Pizzio R, Nabinger C. Native Grasslands at the Core: a New Paradigm of Intensification for the Campos of Southern South America to increase Economic and Environmental Sustainability. Front Sustain Food Syst [Internet]. 2021 [cited 2021 Jul 07];5:547834. Available from: https://bit.ly/3yxt4DQ.
Notes
Author notes
rreyno@inia.org.uy



