
Detección de los genes de β-lactamasas blaTEM, blaSHV y blaCTX-M en aislamientos de Escherichia coli comunitarios
Detection of β-lactamase genes blaTEM, blaSHV and blaCTX-M in community isolates of Escherichia coli
Revista Científica
Universidad de San Carlos de Guatemala, Guatemala
ISSN-e: 2224-5545
Periodicidad: Semestral
vol. 28, núm. 2, 2019
Recepción: 13/02/19
Aprobación: 04 Julio 2019

Resumen: A nivel mundial la resistencia a los antibióticos es un problema de salud pública, tanto en el ámbito hospitalario como en el comunitario. La producción de β-lactamasas es el principal mecanismo de resistencia en enterobacterias y la mayoría de enzimas responsables pertenecen a las familias TEM, SHV y CTX-M. El objetivo de este estudio fue detectar los genes de β-lactamasas blaTEM, blaSHV y blaCTX-M en cepas comunitarias de Escherichia coli productoras de BLEE aisladas de urocultivos de pacientes que acudieron al Laboratorio Clínico Popular de la Universidad de San Carlos de Guatemala en el año 2016. Se detectó la presencia de al menos uno de los genes en el 90% de los 79 aislamientos y un 53.2% presentó los tres genes. La frecuencia fue de 57% para blaCTX-M, 84% para blaSHV y 85% para blaTEM. La detección de los genes codificadores de las enzimas TEM-1, SHV-11, CTX-M15 y CTX-M55 corresponde a la primera caracterización molecular de aislamientos de E. coli productoras de BLEE en Guatemala y son importantes para entender su propagación en el ámbito comunitario. Los aislamientos de E. coli productoras de BLEE mostraron alta resistencia a ciprofloxacina y trimetoprim sulfametoxazol (78%) y bajos niveles de resistencia para fosfomicina (2.5%) y nitrofurantoina (7.6%). El 11.39% de las cepas presentó resistencia a un grupo de antibióticos no betalactámicos. Es importante establecer una vigilancia activa para la resistencia de estos antibióticos en cepas comunitarias ya que son la primera opción de tratamiento para cepas productoras de BLEE.
Palabras clave: β-lactamasas de espectro extendido (BLEE), aislamientos comunitarios, Escherichia coli, TEM, SHV , CTX - M.
Abstract: Globally, resistance to antibiotics is a public health problem, both in the hospital and in the community environment. The production of β-lactamases is the main mechanism of resistance in enterobacteria and usually the cause of resistance are the enzymes to the families TEM, SHV and CTX-M. The principal aim of this study was to detect - β-lactamase genes blaTEM, blaSHV and blaCTX-M in community strains of ESBL-producing Escherichia coli isolated from urine cultures of patients attended in the Laboratorio Clínico Popular of the Universidad de San Carlos de Guatemala in 2016. At least, one of the genes was detected in 90% of the 79 isolates and in 53.2% of the isolates the three genes were detected. The frequency was 57% for blaCTX-M, 84% for blaSHV and 85% for blaTEM. The detection of the genes coding for TEM-1, SHV-11, CTX-M15 and CTX-M55 represent the first molecular characterization of ESBL-producing E. coli isolates in Guatemala and it is important to understand the spread of these strains at the community environment. The ESBL-producing E. coli isolates showed high resistance to ciprofloxacin and trimethoprim sulfamethoxazole (78%) and low resistance levels for fosfomycin (2.5%) and nitrofurantoin (7.6%). The 11.39% of the strains showed resistance to a group of non-beta-lactam antibiotics. It is important to establish an active surveillance for these antibiotics in community strains because they are the first line treatment for strains producing ESBL.
Keywords: extended spectrum β-lactamases (ESBL), community isolates, Escherichia coli, TEM, SHV , CTX - M.
Introducción
La producción de b-lactamasas es el principal mecanismo por el cual se produce la resistencia en bacilos Gram negativo; sobre todo porque los genes que codifican estas enzimas se encuentran en plásmidos que se transmiten a otras bacterias y codifican de forma simultánea genes de resistencia para diversos antibióticos (Geser, Stephan, & Hächler, 2012; Mulvey et al., 2004; Paterson, et al, 2003).
Las β-lactamasas de espectro extendido (BLEE) son enzimas que hidrolizan penicilinas, aztreonam y cefalosporinas que contienen una cadena lateral oximino (primera, segunda y tercera generación). Sin embargo, no actúan sobre las cefamicinas ni carbapenemes, y son inhibidas por el ácido clavulánico y otros inhibidores como el sulbactam o tazobactam (Jeong et al., 2004; Kiratisin, Apisarnthanarak, Laesripa, & Saifon, 2008; Tankhiwale, Jalgaonkar, Ahamad, & Hassani, 2004)
La primera BLEE fue aislada en Alemania en 1983 y desde entonces se han caracterizado más de 200 tipos diferentes en todo el mundo. Las BLEE más diseminadas son las derivadas de las b-lactamasas tipo TEM, SHV y CTX-M por mutaciones puntuales en los genes blaTEM, blaSHV y blaCTX-M que alteran la secuencia de aminoácidos del sitio activo de la enzima y aumentan su capacidad de hidrolisis para ciertos sustratos, como las cefalosporinas de tercera y cuarta generación (Mulvey et al., 2004; Paterson, et al., 2003).
Durante mucho tiempo, las infecciones por cepas productoras de BLEE fueron asociadas a diferentes factores de origen hospitalario que incrementaban la probabilidad de adquirir una infección, como estancias prolongadas en las unidades de cuidados críticos en los hospitales, procedimientos médicos invasivos de larga duración (catéter central, sonda urinaria, tubos orotraqueales y otros), hemodiálisis y malnutrición. Recientemente, diferentes estudios en aislamientos comunitarios de enterobacterias han demostrado que pacientes ambulatorios pueden actuar como reservorios de cepas productoras de BLEE, favoreciendo su diseminación, cuando han tenido infecciones urinarias recurrentes, ingresos previos a hospitales o poseen una afección renal crónica, pulmonar, hepática, cardiovascular o diabetes (Paterson & Bonomo, 2005; Ben-Ami et al., 2006)
Diversos estudios que han caracterizado molecularmente las BLEE de aislamientos hospitalarios y comunitarios, determinaron una alta frecuencia de b-lactamasas derivadas de TEM y SHV. Sin embargo, se ha producido un incremento en la frecuencia de cepas productoras de BLEE de la clase CTX-M, que se ha relacionado al aumento en el uso de ceftriaxona a nivel mundial. Estos hallazgos son motivo de preocupación por la tendencia mundial de aparición de infecciones adquiridas en la comunidad causadas por cepas productoras de BLEE del tipo CTX-M, cuya propagación es asociada a políticas ineficientes de antibióticos a nivel hospitalario y ambulatorio (Blanco et al., 2013; Mendonça, Leitão, Manageiro, Ferreira, & Caniça, 2007; Paterson & Bonomo, 2005).
En Guatemala se han realizado estudios para estimar la prevalencia de BLEE en varios centros de atención hospitalaria y aislamientos provenientes de la comunidad reportando prevalencias de 31.9% y 27%, respectivamente (Alvarado & Mejía, 2014; Gordillo, Mejia, & Matheu, 2014). Sin embargo, no existen datos respecto al tipo específico de BLEE producido por estas enterobacterias, sobre todo en los aislamientos comunitarios.
El presente estudio tuvo como finalidad detectar los genes de β-lactamasas blaTEM, blaSHV y blaCTX-M en aislamientos comunitarios de Escherichia coli a partir de urocultivos de pacientes atendidos en el Laboratorio Clínico Popular (LABOCLIP) de la Universidad de San Carlos de Guatemala, durante el primer cuatrimestre del año 2016. Para determinar los genes se utilizó la reacción en cadena de la polimerasa (PCR) y secuenciación de Sanger bidireccional. Los resultados obtenidos son los primeros datos a nivel nacional sobre la epidemiología molecular de las cepas de Escherichia coli productoras de BLEE que circulan a nivel comunitario en el país, lo cual es de importancia para el monitoreo y vigilancia de la resistencia antimicrobiana.
Materiales y métodos
Selección de las cepas
Durante los meses de enero a abril del año 2016 se recolectaron 94 aislamientos comunitarios de E. coli productores de BLEE a partir de urocultivos en el área de Microbiología del Laboratorio Clínico Popular (LABOCLIP). El mecanismo de resistencia fue detectado a través del método de Kirby-Bauer con discos de 30 µg de cefotaxime (OXOID, Hampshire UK), 30 µg de ceftazidime (OXOID, Hampshire UK) y 20/10 µg de amoxicilina/ácido clavulánico (OXOID, Hampshire UK). Quince cultivos fueron descartados debido a que, no se obtuvo crecimiento bacteriano y la calidad o cantidad del DNA extraído no fue adecuada luego de dos repeticiones.
Enriquecimiento y extracción de DNA
Las cepas de E. coli fueron inoculadas en 5 mL de caldo LB (Luria Berthani) para su enriquecimiento por 48 h a 37 °C. Posteriormente se tomaron 400 µL del cultivo para realizar la extracción de DNA por medio del kit comercial Wizard® Genomic DNA Purification según las instrucciones del fabricante (Promega, Madison USA).
Reacción en cadena de la polimerasa (PCR) de los genes de β-lactamasas
Secuenciación de Sanger
Los productos de PCR obtenidos para cinco cepas fueron secuenciados por el método de Sanger bidireccional en la empresa Macrogen, USA (Maryland, USA). Los resultados fueron analizados en el programa Geneious R8.8.1.8. Las secuencias fueron traducidas a proteínas y analizadas en el software CLC sequence viewer 8.0 Qiagen®.
Análisis de resultados
Se utilizaron frecuencias, mediana y proporciones para describir el sexo y edad de los pacientes de quienes provenían los aislamientos, presencia o ausencia de los genes de β-lactamasas y la resistencia o susceptibilidad a antibióticos. Se buscaron diferencias entre la resistencia a los antibióticos no β-lactámicos de los aislamientos provenientes de pacientes con y sin historia de enfermedades del tracto urinario. La estadística descriptiva de los aislamientos y los perfiles de resistencia fueron analizados en el programa Microsoft Excel. Las comparaciones entre grupos se realizaron con la prueba Ji2 a través de la herramienta informática OpenEpi versión 3.01, y un valor de p ≤ .05.
Resultados
El 84.0% (66/79) de los aislamientos procedían de urocultivos de mujeres. La mediana de la edad fue de 57 años, rangos intercuartilico (RIC= 26 - 67 años) para todos los pacientes, y 31.6% (25/79) presentaba historia de infecciones del tracto urinario (ITU) recurrentes.
Por medio de la técnica de reacción en cadena de la polimerasa (PCR) se analizaron 79 cepas productoras de BLEE y se determinó la presencia de los genes blaTEM, blaSHV y blaCTX-M (Figura 1). En 71 (90%) aislamientos se detectó la presencia de, al menos, uno de los genes analizados.
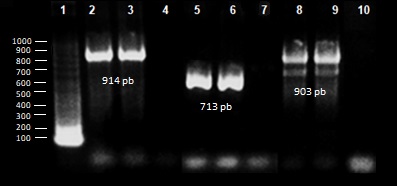
El 82.3% (65/79) de las cepas presentó más de uno de los tipos de β-lactamasas, y en más de la mitad se detectó la presencia de los tres genes. (53.2%). Las genes codificantes para las enzimas de tipo TEM y SHV fueron detectados de forma única en 3.8% de los aislamientos. Por el contrario, los genes codificantes de las enzimas de tipo CTX-M fueron detectados siempre de forma conjunta con los de las enzimas TEM y/o SHV. En 10.1% de los aislamientos no se detectó la presencia de ninguno de los genes de interés (Tabla 1).
| Genes de β-lactamasas | Aislamientos (N=79) | % de aislamientos |
| blaTEM, blaSHV y blaCTX-M | 42 | 53.2 |
| blaSHV, blaCTX-M | 2 | 2.5 |
| blaTEM, blaCTX-M | 1 | 1.3 |
| blaTEM,blaSHV | 20 | 25.3 |
| blaTEM | 3 | 3.8 |
| blaSHV | 3 | 3.8 |
| Negativo (otra BLEE) | 8 | 10.1 |
| Cepa analizada | Tipo de β-lactamasa |
| ECO-57 | CTX-M15, SHV-11 |
| ECO-96 | CTX-M55 |
| ECO-97 | TEM-1 |
| ECO-109 | CTX-M15, SHV-11 yTEM-1 |
| ECO-112 | SHV-11 yTEM-1 |
La Tabla 3 muestra el perfil de resistencia de las cepas evaluadas frente a los antibióticos no betalactámicos, incluidos en el antibiograma de forma rutinaria en el LABOCLIP. Los porcentajes más altos de resistencia corresponden a ciprofloxacina y trimetoprim sulfametoxazol (ambos 62/79). Por el contrario, la resistencia fue baja para fosfomicina (2/79) y nitrofurantoina (6/79).
| Antibiótico | Resistenteα | Susceptible | ||
| Fosfomicinaβ | 2 | 2.5% | 68 | 86.1% |
| Nitrofurantoina | 6 | 7.6% | 73 | 92.4% |
| Ciprofloxacina | 62 | 78.5% | 17 | 21.5% |
| Amoxicilina/Ácido clavulánico | 34 | 43.0% | 45 | 57.0% |
| Trimetoprim sulfametoxazol | 62 | 78.5% | 17 | 21.5% |
| α Incluye resultados resistente e intermedio del antibiograma βNueve de los aislamientos no fueron evaluados para fosfomicina | ||||
El 11.39% (9/79) de las cepas estudiadas presentó resistencia a un antibiótico no betalactámico, y en la mayoría la resistencia fue múltiple. El 59.49 % (47/79) fue resistente a ciprofloxacina y trimetoprim sulfametoxazol de forma combinada, siendo este perfil de resistencia el más común. Además, 29.11% de las cepas con resistencia a ambos antibióticos presentó también resistencia a la combinación de amoxicilina/ácido clavulánico.
No se encontró diferencia significativa entre la resistencia a ciprofloxacina, trimetoprim sulfametoxazol y amoxicilina/ácido clavulánico cuando se compararon a los pacientes con historia de ITU recurrentes y los que no tenían este antecedente (p = .84, p = .17 y p = .55, respectivamente).
Discusión
El LABOCLIP es un laboratorio escuela que atiende diariamente aproximadamente 300 pacientes ambulatorios y la mayoría ellos tiene recursos limitados. Según el registro del área de Microbiología Clínica, en el año 2016 en el 36% de los urocultivos se aisló E. coli BLEE positivo y esta bacteria fue el patógeno más frecuente asociado a infecciones del tracto urinario con 93%.
A nivel mundial se ha reportado un incremento alarmante en las cepas productoras de BLEE de aislamientos comunitarios; sobre todo en patógenos de la familia Enterobacteriaceae (Fan et al., 2014; Pitout, Nordmann, Laupland & Poirel, 2005; Shaikh, Fatima, Shakil, & Rizvi, 2015; Tumbarello et al., 2006). La detección y caracterización de estos aislamientos es de vital importancia para la selección de terapia, control y prevención de los mismos. Por ello, se planteó la detección de los genes blaTEM, blaSHV y blaCTX-M en aislamientos comunitarios de E. coli productores de BLEE aislados de urocultivos.
Se detectó la presencia de, al menos, uno de los genes analizados en 89.8% de los aislamientos (71/79). Los genes blaTEM y blaSHV fueron los más frecuentes con un 84 y 85%, respectivamente. Sin embargo, uno de los principales problemas de la caracterización de las BLEE es que frecuentemente se asocian a β-lactamasas de amplio espectro (BLEA), sobre todo de los tipos TEM y SHV de las cuales se originan, lo cual hace necesario, la identificación del tipo específico de enzima para poder discriminar entre unas y otras (Bradford, 2001; Jacoby & Munoz-Prince, 2005; Paterson & Bonomo, 2005).
Por el contrario, la detección del gen blaCTX-M en aislamientos productores de BLEE es evidencia suficiente para explicar el fenotipo resistente. Es decir que, en el 57% de los aislamientos analizados, la enzima CTX-M es responsable del patrón de resistencia mostrado, independientemente de la presencia de las enzimas del tipo TEM y SHV, cuyos genes también fueron detectados. Este porcentaje concuerda con estudios realizados en cepas E. coli comunitarias a nivel mundial, en donde se reportan entre 40 y 70% de BLEE tipo CTX-M (Brigante et al., 2005; Rodriguez-Villalobos et al., 2011; Shaikh et al., 2015).
En el 10.1 % de los aislamientos en los que no se detectó la presencia de los genes analizados, es indispensable evaluar la presencia de genes que codifican para otros tipos de enzimas como GES, BES, TLA, SFO e IBC que presentan un espectro extendido y han sido descritas en enterobacterias (Bradford, 2001; Paterson & Bonomo, 2005; Shaikh et al., 2015).
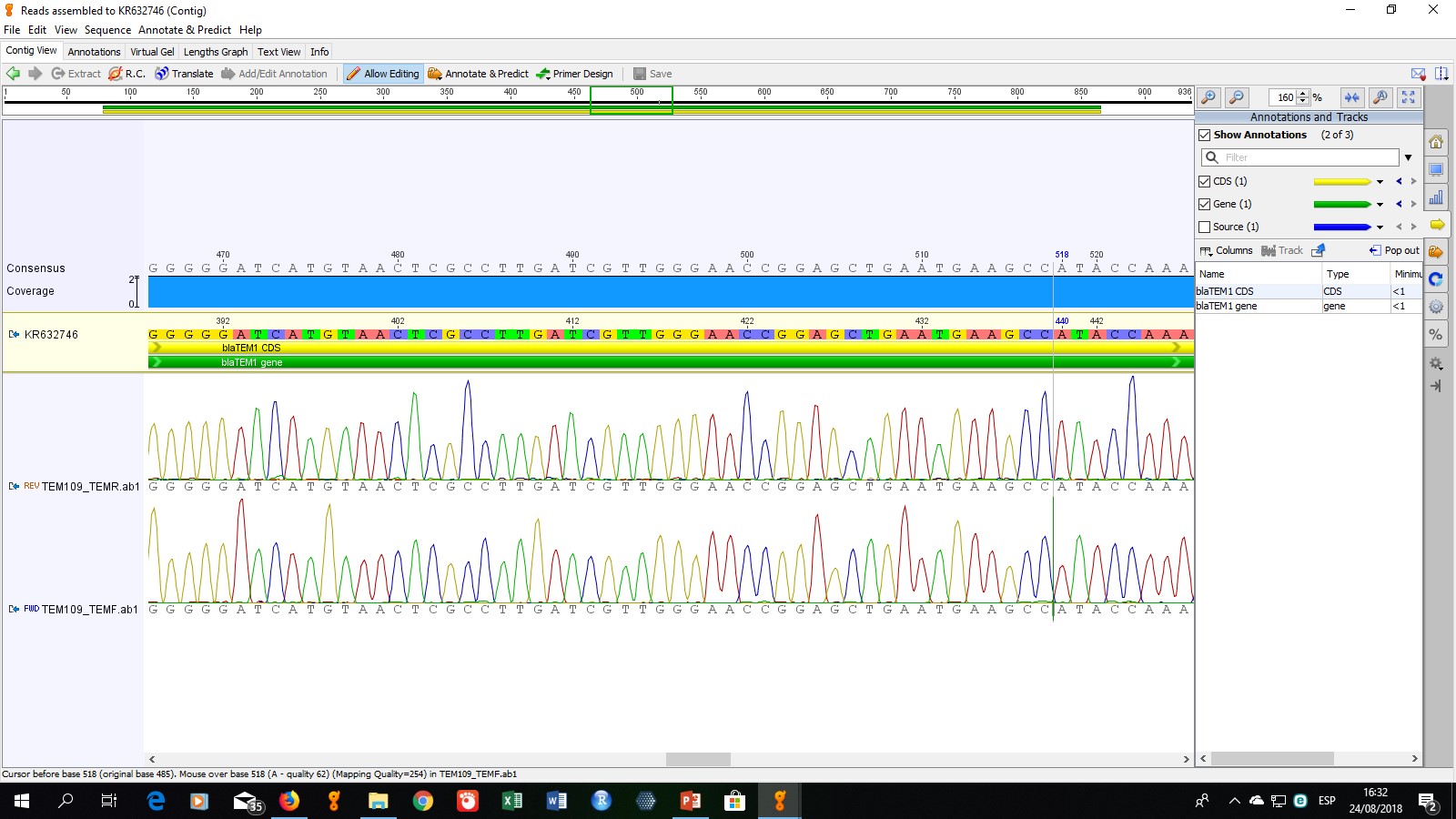
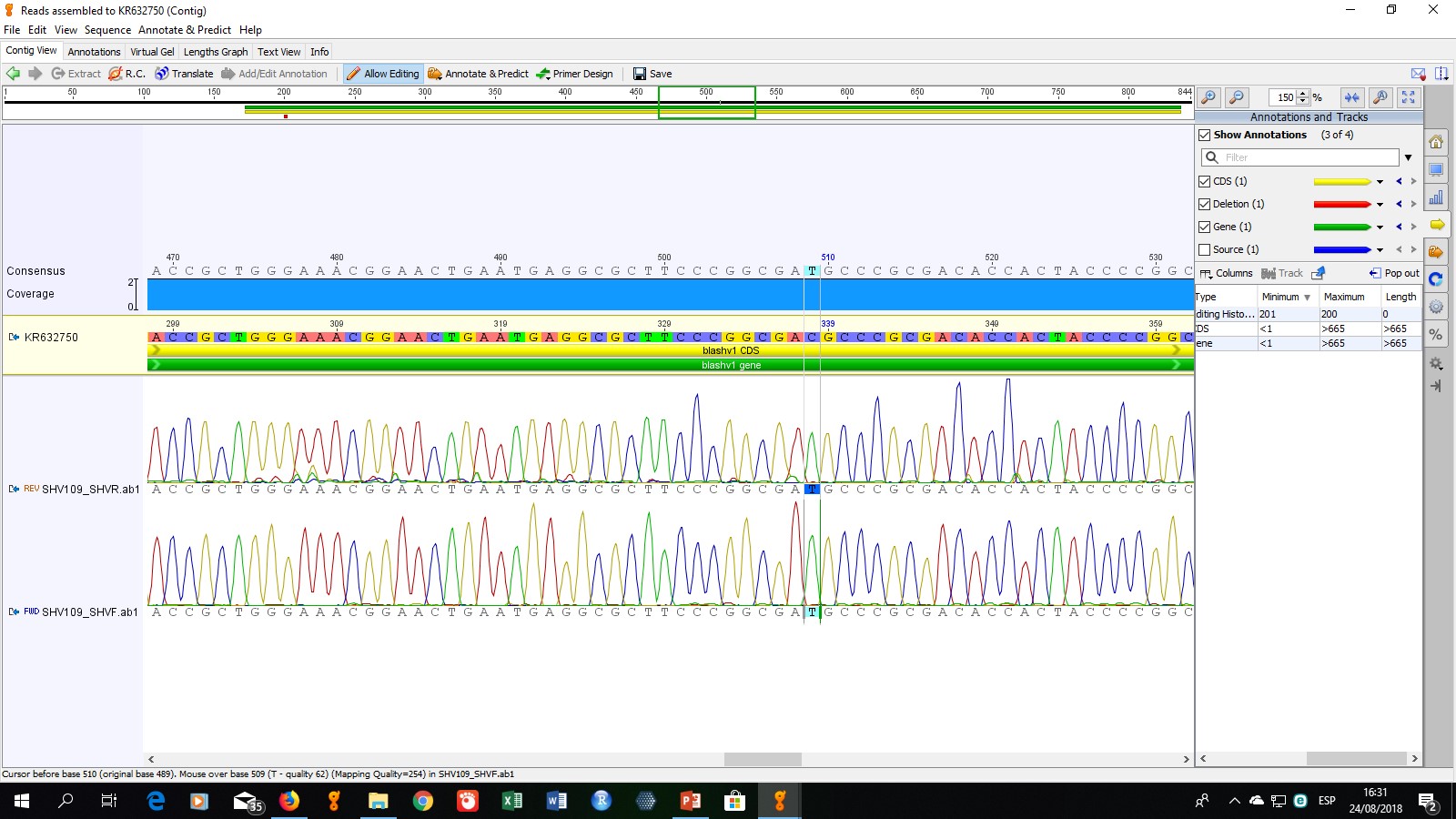
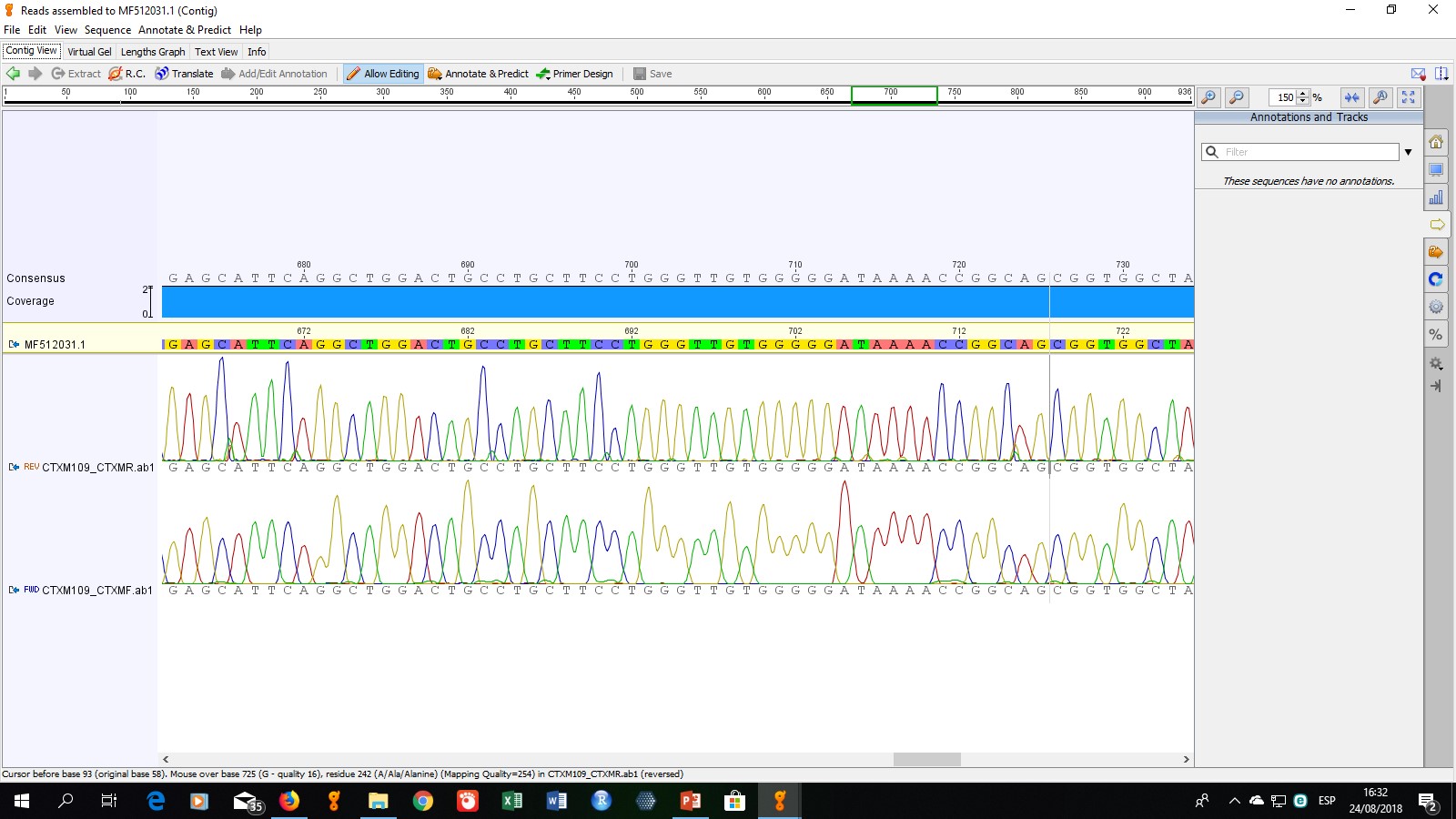
Para determinar el tipo específico de enzimas presentes, se seleccionaron cinco cepas para la secuenciación de los productos de PCR. La cepa codificada como ECO-109 fue seleccionada debido a que mostró un perfil de multiresistencia (resistente a todos los antibióticos no betalactámicos evaluados), las cepas ECO-57, ECO-96, ECO-97 y ECO-112 fueron seleccionadas aleatoriamente. Los resultados de secuenciación obtenidos mostraron que los genes presentes, corresponden a las enzimas TEM-1, SHV-11, CTX-M15 y CTX-M55.
La enzima TEM-1 es una β-lactamasa de espectro ampliado identificada por primera vez en 1965 y es responsable de, aproximadamente, el 90% de la resistencia a ampicilina mediada por plásmidos en E. coli (Bradford, 2001; Paterson & Bonomo, 2005). Esta enzima fue detectada en las cepas ECO-97, ECO-109 y ECO-112, por lo que en estos tres aislamientos se tiene la certeza de que el fenotipo BLEE mostrado no se debe a enzimas del tipo TEM.
La enzima SHV-11, cuyo gen fue detectado en las tres cepas secuenciadas (ECO-57, ECO-109 y ECO-112) fue descrita en 1997 y se diferencia de la enzima SHV-1 (la primera enzima de este grupo descrita) solamente por el cambio de una leucina por una glutamina en la posición 35. Este cambio es distante del sitio activo de la enzima, por lo que el espectro de hidrólisis de SHV-11 es igual a SHV-1 (Nüesch-Inderbinen, Kayser, & Hächler, 1997). Por tanto, la presencia de esta enzima, al igual que la presencia de TEM-1, nos indica que el fenotipo BLEE determinado en las cepas es causado por otra enzima.
La enzima CTX-M15, cuyo gen fue detectado en dos aislamientos de este estudio, fue aislada por primera vez en India y es la enzima del tipo CTX-M más diseminada a nivel mundial. Forma parte de la denominada “pandemia CTX-M” asociada a infecciones del tracto urinario (ITU) y bacteremias adquiridas en la comunidad (Coque et al., 2008; Hijazi, Fawzi, Ali, & Abd El Galil, 2016; Moubareck et al., 2005). El gen blaCTX-M15 se ha encontrado en elementos móviles altamente eficaces como los plásmidos IncFI, IncFII, IncHI2, IncI, IncN, IncP-1-a, IncL/M e IncA/C; y las secuencias ISEcp1 e ISCR1 (Brigante et al., 2005; Peirano & Pitout, 2010). Recientes estudios de vigilancia informan altos porcentajes de aislamientos de E. coli productores de CTX-M15 en Europa, Asia y América del Sur (Hijazi et al., 2016; Lau et al., 2008; Shi et al., 2015).
Estudios de epidemiología molecular con la técnica de MLST (multilocus sequencing typing) sugieren que la rápida propagación de E. coli productora de CTX-M15 a nivel mundial se debe al clon ST131. Este clon altamente virulento contiene además múltiples genes de resistencia como blaTEM-1, blaOXA-1, aaa(6’)-Ib-cr, tetA y aaa(3)-II, contenidos en integrones de clase 1 o asociados al plásmido IncFII (Machado et al., 2006; Peirano, Costello, & Pitout, 2010; Shi et al., 2015). Estudios futuros de clonalidad en cepas de E. coli productoras de BLEE, podrían explicar la alta frecuencia de cepas co-productoras de enzimas del tipo TEM-1 y CTX-M-15 (54%) obtenidas en este estudio, así como, la resistencia combinada a flouroquinolonas.
Las enzimas aaa(3)-II y aaa(6’)-Ib-cr se han asociado a E. coli productoras de CTX-M15 (Peirano & Pitout, 2010; Xiao & Hu, 2012). Estás enzimas modifican los aminoglucósidos y los inactivan. En este estudio no fue posible determinar la resistencia a este grupo de antibióticos, debido a que no son evaluados de forma rutinaria en el antibiograma utilizado para Gram negativos en el LABOCLIP. La cepa ECO-109 fue resistente a los antibióticos de rutina evaluados, por lo que se enfrentó a dos aminoglucósidos, y mostró resistencia frente a gentamicina y susceptibilidad a amikacina. En esta cepa particular, podría analizarse la presencia de estos dos genes de resistencia para confirmar el fenotipo mostrado.
La enzima aaa(6’)-Ib-cr tiene también la capacidad de acetilar, y con ello inactivar, fluoroquinolonas que presentan el nitrógeno amino libre en el anillo de piperazina (norfloxacina y ciprofloxacina) (Park, Robicsek, Jacoby, Sahm, & Hooper, 2006; Peirano & Pitout, 2010). La presencia combinada de esta enzima con blaCTX-M15 explicaría la elevada resistencia a fluoroquinolonas (ciprofloxacina) que mostraron las cepas analizadas (78.5%), aunque debe evaluarse la presencia de otros genes como gyrA, parC (cromosómicos), qnrA, qnrB y qnrS (plasmídicos) que también se asocian a resistencia en estos antibióticos (Hernández, 2010; Jacoby et al., 2006; Peirano et al., 2010; Peirano & Pitout, 2010). La elevada tasa de resistencia a fluoroquinolonas en cepas productoras de BLEE encontrada, concuerda con prevalencias reportadas en otros estudios como el de Rodríguez-Baño y colaboradores que encontraron 77% de resistencia a fluoroquinolonas en cepas productoras de BLEE (Coque et al., 2008; García-Gómez, Guio, Hernández, Vilar & Pijoán, 2015; Rodríguez-Baño et al., 2004; Rodriguez-Villalobos et al., 2011).
La enzima CTX-M55 pertenece al subgrupo CTX-M1 al igual que la enzima CTX-M15, de la cual se diferencia simplemente por tener una valina en vez de alanina en la posición 77, por lo que se cree que se ha derivado de esta enzima. Fue descrita por primera vez en Tailandia y estudios estructurales han mostrado que al igual que CTX-M15 se asocia a la presencia de ISEcp1 (Kiratisin, et al., 2007). En China, esta es la segunda enzima del tipo CTX-M más frecuente y ha sido aislada en humanos y animales (Kim et al., 2017). La cepa ECO-96 que mostró la presencia de esta enzima fue resistente a fosfomicina, un dato de interés, sobre todo porque se ha descrito la producción conjunta de la enzima CTX-M55 y el gen fosA3 en aislamientos de E. coli en Brasil y Francia (Cunha, Lincopan, Cerdeira, & Esposito, 2017; Fernandes, Sellera, Moura, Souza, & Lincopan, 2018; Lupo, Saras, Madec, & Haenni, 2018).
La nitrofurantoína y la fosfomicina fueron los antibióticos que presentaron menor tasa de resistencia (7.6 y 2.5 % respectivamente). Sin embargo, éstas son elevadas en comparación con reportes internacionales previos de 0.3 % de cepas resistentes a fosfomicina y 1.5 % a nitrofurantoína en aislamientos comunitarios. Los niveles elevados de resistencia a antibióticos se asocian políticas ineficientes para el manejo de antibióticos a nivel hospitalario y ambulatorio, déficit sanitario, presencia y propagación conjunta de genes de resistencia a varios tipos de antibióticos y la colonización de reservorios animales y humanos no identificados. (Dash, Al-zarouni, Al-kous, & Shehhi, 2008; Zhanel et al., 2006).
El elevado número de organismos multirresistentes en pacientes ambulatorios es una amenaza emergente para la salud pública en el país, pues evidencia la circulación de cepas bacterianas resistentes a nivel comunitario. La resistencia mostrada a fluoroquinolonas, trimetroprim sulfametoxazole y amoxicilina/ácido clavulánico en cepas productoras de BLEE limita la terapia al uso de carbapenemes, nitrofurantoína y fosfomicina como la primera opción para el tratamiento de infecciones del tracto urinario que se originan en la comunidad. Ello es alarmante, ya que en ámbitos hospitalarios se ha reportado una frecuencia de 91% de cepas portadoras de genes de resistencia a carbapenemes (Velásquez Porta, 2016).
Agradecimientos
En especial al área de Microbiología del Laboratorio Clínico Popular por su valiosa colaboración para la realización de esta investigación. A INVEGEM/Rozas Botrán por la formación profesional y el apoyo brindado, en especial su directora científica y al personal del laboratorio de Biología Molecular.
Referencias
Alvarado, J., & Mejía, C. (2014). Resistencia Bacteriana en Infecciones del Tracto Urinario de Origen Comunitario. Revista de Medicina Interna de Guatemala, 20, 24-30.
Ben-Ami, R., Schwaber, M. J., Navon-Venezia, S., Schwartz, D., Giladi, M., Chmelnitsky, I., ... & Carmeli, Y. (2006). Influx of Extended spectrum β-lactamase producing Enterobacteriaceae into the Hospital. Clinical Infectious Diseases, 42(7), 925-934.
Blanco, J., Mora, A., Mamani, R., López, C., Blanco, M., Dahbi, G. & Rodríguez-Baño, J. (2013). Four main virotypes among extended-spectrum-β-lactamase-producing isolates of Escherichia coli O25b:H4-B2-ST131: Bacterial, epidemiological, and clinical characteristics. Journal of Clinical Microbiology, 51(10), 3358–3367.
Bradford, P. A. (2001). Extended-Spectrum β-Lactamases in the 21st Century : Characterization, Epidemiology , and Detection of This Important Resistance Threat. Clinical Microbiology Reviews, 14(4), 933-951.
Brigante, G., Luzzaro, F., Perilli, M., Lombardi, G., Colì, A., Rossolini, G. M., … Toniolo, A. (2005). Evolution of CTX-M-type β-lactamases in isolates of Escherichia coli infecting hospital and community patients. International Journal of Antimicrobial Agents, 25(2), 157-162.
Coque, T. M., Novais, Â., Carattoli, A., Poirel, L., Pitout, J., Peixe, L., … Nordmann, P. (2008). Dissemination of clonally related Escherichia coli strains expressing extended-spectrum β-lactamase CTX-M-15. Emerging Infectious Diseases, 14(2), 195-200.
Cunha, M. P. V., Lincopan, N., Cerdeira, L., & Esposito, F. (2017). Coexistence of CTX-M-2, CTX-M-55, CMY-2, FosA3, and QnrB19 in Extraintestinal Pathogenic Escherichia coli from Poultry in Brazil. Antimicrobial Agents and Chemotherapy Downloaded, 61(4), 1-3.
Dash, N., Al-zarouni, M., Al-kous, N., & Shehhi, F. A. (2008). Distribution and Resistance Trends of Community Associated Urinary Tract Pathogens in Sharjah, UAE. Microbiology Insights, 3, 41-45.
Fan, N. C., Chen, H. H., Chen, C. L., Ou, L. S., Lin, T. Y., Tsai, M. H., & Chiu, C. H. (2014). Rise of community-onset urinary tract infection caused by extended-spectrum β-lactamase-producing Escherichia coli in children. Journal of Microbiology, Immunology and Infection, 47(5), 399-405.
Fernandes, M. R., Sellera, F. P., Moura, Q., Souza, T. A., & Lincopan, N. (2018). Draft genome sequence of a CTX-M-8, CTX-M-55 and FosA3 co-producing Escherichia coli ST117/B2 isolated from an asymptomatic carrier. Journal of Global Antimicrobial Resistance, 12, 183-184.
García-Gómez, M., Guío, L., Hernández, J. L., Vilar, B., & Pijoán, J. I. (2015). Original Bacteriemias por enterobacterias productoras de beta-lactamasas (BLEE , AmpC y carbapenemasas): asociación con los cuidados sanitarios y los pacientes oncológicos. Revista Española de Quimioterapia, 28(5), 256-262.
Geser, N., Stephan, R., & Hächler, H. (2012). Occurrence and characteristics of extended-spectrum β-lactamase (ESBL) producing Enterobacteriaceae in food producing animals, minced meat and raw milk. BMC Veterinary Research, 8(1), 21.
Gordillo, M. R., Mejia, C. R., & Matheu, J. (2014). Presencia de β-lactamasas de expectro extendido, BLEE, y enterobacterias portadoras de carbapenemasas KPC, en enterobacterias en el Hospital Roosevelt, 2011 Y 2012. Revista de Medicina Interna de Guatemala, 17(9), 8–22.
Hernández, E. (2010). Escherichia coli productores de BLEE aislados de urocultivo: implicaciones en el dignóstico y tratamiento de la infección urinaria. Madrid: Universidad Complutense de Madrid.
Hijazi, S. M., Fawzi, M. A., Ali, F. M., & Abd El Galil, K. H. (2016). Prevalence and characterization of extended-spectrum beta-lactamases producing Enterobacteriaceae in healthy children and associated risk factors. Annals of Clinical Microbiology and Antimicrobials, 15(1), 1-9.
Jacoby, G. A., & Munoz-Prince, L. S. (2005). Mechanisms of Disease: The new β-Lactamases. The New. England. Journal of Medicine, 352(4), 380–391.
Jacoby, G. A, Walsh, K. E., Mills, D. M., Walker, J., Oh, H., Robicsek, A. & Hooper, D. C. (2006). qnrB, Another Plasmid-Mediated Gene for Quinolone Resistance. Antimicrobial Agents and Chemotherapy, 50(4), 1178–1182.
Jeong, S. H., Bae, I. K., Lee, J. H., Sohn, S. G., Kang, G. H., Jeon, G. J.& Lee, S. H. (2004). Molecular Characterization of Extended-Spectrum Beta-Lactamases Produced by Clinical Isolates of Klebsiella pneumoniae and Escherichia coli from a Korean Nationwide Survey. Journal of clinical microbiology, 42(7), 2902–2906.
Kim, J. S., Kim, S., Park, J., Shin, E., Yun, Y. S., Lee, D. Y., … Kim, J. (2017). Plasmid-mediated transfer of CTX-M-55 extended-spectrum beta-lactamase among different strains of Salmonella and Shigella spp. in the Republic of Korea. Diagnostic Microbiology and Infectious Disease, 89(1), 86–88.
Kiratisin, P., Apisarnthanarak, A., Laesripa, C., & Saifon, P. (2008). Molecular characterization and epidemiology of extended spectrum β-lactamase producing Escherichia coli and Klebsiella pneumoniae isolates causing health care-associated infection in Thailand, where the CTX-M family is endemic. Antimicrobial Agents and Chemotherapy, 52(8), 2818–2824.
Kiratisin, P., Apisarnthanarak, A., Saifon, P., Laesripa, C., Kitphati, R., & Mundy, L. M. (2007). The emergence of a novel ceftazidime-resistant CTX-M extended-spectrum β-lactamase, CTX-M-55, in both community-onset and hospital-acquired infections in Thailand. Diagnostic Microbiology and Infectious Disease, 58(3), 349–355.
Lau, S. H., Kaufmann, M. E., Livermore, D. M., Woodford, N., Willshaw, G. A., Cheasty, T., … Upton, M. (2008). UK epidemic Escherichia coli strains A-E, with CTX-M-15 β-lactamase, all belong to the international O25:H4-ST131 clone. Journal of Antimicrobial Chemotherapy, 62(6), 1241–1244.
Lupo, A., Saras, E., Madec, J. Y., & Haenni, M. (2018). Emergence of blaCTX-M-55 associated with fosA, rmtB and mcr gene variants in Escherichia coli from various animal species in France. Journal of Antimicrobial Chemotherapy, 73(4), 867–872.
Machado, E., Coque, T. M., Cantón, R., Baquero, F., Sousa, J. C., & Peixe, L. (2006). Dissemination in Portugal of CTX-M15, OXA-1-, and TEM-1-producing Enterobacteriaceae strains containing the aac(6′)-Ib-cr gene, which encodes an aminoglycoside- and fluoroquinolone-modifying enzyme. Antimicrobial Agents and Chemotherapy, 50(9), 3220–3221.
Mendonça, N., Leitão, J., Manageiro, V., Ferreira, E., & Caniça, M. (2007). Spread of extended-spectrum β-lactamase CTX-M-producing Escherichia coli clinical isolates in community and nosocomial environments in Portugal. Antimicrobial Agents and Chemotherapy, 51(6), 1946–1955.
Moubareck, C., Daoud, Z., Hakimé, N. I., Hamzé, M., Mangeney, N., Matta, H., … Doucet-Populaire, F. (2005). Countrywide spread of community and hospital acquired extended-spectrum β-lactamase CTX-M15 producing Enterobacteriaceae in Lebanon. Journal of Clinical Microbiology, 43(7), 3309–3313.
Mulvey, M. R., Bryce, E., Boyd, D., Ofner-agostini, M., Christianson, S., Simor, A. E., … Program, I. S. (2004). Ambler Class A Extended Spectrum Beta-Lactamase Producing. Society, 48(4), 1204–1214.
Nüesch-Inderbinen, M. T., Kayser, F. H., & Hächler, H. (1997). Survey and molecular genetics of SHV beta-lactamases in Enterobacteriaceae in Switzerland: Two novel enzymes, SHV-11 and SHV-12. Antimicrobial Agents and Chemotherapy, 41(5), 943–949.
Park, C. H., Robicsek, A., Jacoby, G. A., Sahm, D., & Hooper, D. C. (2006). Prevalence in the United States of aac(6′)-Ib-cr encoding a ciprofloxacin-modifying enzyme. Antimicrobial Agents and Chemotherapy, 50(11), 3953–3955.
Paterson, D. L., & Bonomo, R. (2005). Extended-Spectrum β-Lactamases : a Clinical Update. Society, 18(4), 657–686.
Paterson, D. L., Hujer, K. M., Hujer, A. M., Yeiser, B., Bonomo, M. D., Rice, L. B., & Bonomo, R. (2003). Extended-Spectrum Beta-Lactamases in Klebsiella pneumoniae Bloodstream Isolates from Seven Countries: Dominance and Widespread Prevalence of SHV- and CTX-M-Type β-Lactamases. Antimicrobial Agents and Chemotherapy, 47(11), 3554–3560.
Peirano, G., Costello, M., & Pitout, J. D. D. (2010). Molecular characteristics of extended-spectrum β-lactamase-producing Escherichia coli from the Chicago area: High prevalence of ST131 producing CTX-M15 in community hospitals. International Journal of Antimicrobial Agents, 36(1), 19–23.
Peirano, G., & Pitout, J. D. (2010). Molecular epidemiology of Escherichia coli producing CTX-M β-lactamases: the worldwide emergence of clone ST131 O25:H4. International Journal of Antimicrobial Agents, 35(4), 316–321.
Pitout, J. D., Nordmann, P., Laupland, K. B., & Poirel, L. (2005). Emergence of Enterobacteriaceae producing extended-spectrum β-lactamases (ESBLs) in the community. Journal of Antimicrobial Chemotherapy, 5, 52–59.
Rodríguez-Baño, J., Navarro, M. D., Romero, L., Martínez-Martínez, L., Muniain, M. A., Perea, E. J., … Pascual, A. (2004). Epidemiology and Clinical Features of Infections Caused by Extended-Spectrum Beta-Lactamase-Producing Escherichia coli in Nonhospitalized Patients. Journal of Clinical Microbiology, 42(3), 1089–1094.
Rodriguez-Villalobos, H., Bogaerts, P., Berhin, C., Bauraing, C., Deplano, A., Montesinos, I., … Glupczynski, Y. (2011). Trends in production of extended-spectrum β-lactamases among Enterobacteriaceae of clinical interest: Results of a nationwide survey in Belgian hospitals. Journal of Antimicrobial Chemotherapy, 66(1), 37–47.
Shaikh, S., Fatima, J., Shakil, S., & Rizvi, S. M. D. (2015). Antibiotic resistance and extended spectrum beta-lactamases: Types, epidemiology and treatment. Saudi Journal of Biological Sciences, 22(1), 90–101.
Shi, H., Sun, F., Chen, J., Ou, Q., Feng, W., Yong, X., & Xia, P. (2015). Epidemiology of CTX-M-type extended-spectrum beta-lactamase (ESBL)-producing nosocomial -Escherichia coli infection in China. Annals of Clinical Microbiology and Antimicrobials, 14(1), 1–5.
Tankhiwale, S. S., Jalgaonkar, S. V., Ahamad, S., & Hassani, U. (2004). Evaluation of extended spectrum beta lactamase in urinary isolates. Indian Journal of Medical Research, 120(6), 553–556.
Tumbarello, M., Spanu, T., Sanguinetti, M., Citton, R., Montuori, E., Leone, F., … Cauda, R. (2006). Bloodstream Infections Caused by Extended-Spectrum-Betalactamase-Producing Klebsiella pneumoniae: Risk Factors , Molecular Epidemiology, and Clinical Outcome. American Society for Microbiology, 50(2), 498–504.
Velásquez Porta, T. (2016). Detección de los genes de carbapenemasas blaKPC y blaNDM en aislamientos de Klebsiella pneumoniae del Hospital General San Juan de Dios de la ciudad de Guatemala. (Tesis de maestría). Universidad de San Carlos de Guatemala, Guatemala.
Xiao, Y., & Hu, Y. (2012). The Major Aminoglycoside-Modifying Enzyme AAC(3)-II Found in Escherichia coli Determines a Significant Disparity in Its Resistance to Gentamicin and Amikacin in China. Microbial Drug Resistance, 18(1), 42–46.
Zhanel, G. G., Hisanaga, T. L., Laing, N. M., DeCorby, M. R., Nichol, K. A., Palatnik, L. P., … Hoban, D. J. (2006). Antibiotic resistance in Escherichia coli outpatient urinary isolates: final results from the North American Urinary Tract Infection Collaborative Alliance (Nautica). International Journal of Antimicrobial Agents, 27(6), 468–475.

