2021
101
15062021
06072021
 Thaís Ribeiro Costa thais.costa@ufvjm.edu.br
Thaís Ribeiro Costa thais.costa@ufvjm.edu.br
UFVJM, Brasil
Cristiane Coelho de Moura
UFVJM, Brasil
Evandro Luiz Mendonça Machado
UFVJM, Brasil
Anne Priscila Dias Gonzaga
UFVJM, Brasil
Resumo: Capões abrigam um dos maiores patrimônios biológicos da Reserva da Biosfera da Serra do Espinhaço e sua conservação depende do conhecimento amplo e detalhado de suas espécies. Assim, o objetivo deste estudo foi avaliar a composição florística da comunidade arbórea desta vegetação. Para isso, selecionamos 22 capões em duas Unidades de Conservação: Parque Nacional das Sempre-Vivas e Parque Estadual do Biribiri. A amostragem da vegetação arbórea foi realizada em parcelas permanentes instaladas aleatoriamente no interior das florestas. Para o conjunto total dos capões, encontramos 1692 indivíduos arbóreos, distribuídos em 112 espécies e 40 famílias botânicas. A espécie mais representativa foi Richeria grandis. A composição das áreas compreendeu espécies generalistas e típicas de ambientes úmidos e vegetação de savana (Cerrado). Nossos resultados demostram que apesar de ocuparem pequena porção territorial, estes ambientes constituem importantes centros de biodiversidade multitaxonômica.
Palavras-chave: Florestas úmidas, Matas de galeria inundáveis, Riqueza de espécies, Unidades de Conservação, Preservação.
Abstract: “Capões” are home to one of the greatest biological assets of the Serra do Espinhaço Biosphere Reserve and its conservation depends on extensive and detailed knowledge of its species. Thus, the aim of this study was to evaluate the floristic composition of the arboreal community of this vegetation. For this purpose, we selected 22 capões marshes and forests in two Conservation Units: Semper-Vivas National Park and Biribiri State Park. The sampling of arboreal vegetation was carried out in permanent plots installed randomly inside the forests. For the total set of capões, we found 1692 arboreal individuals, distributed in 112 species and 40 botanical families. The most representative species was Richeria grandis. The composition of the areas included generalist and typical species from humid environments and savanna (Cerrado) vegetation. Our results show that despite occupying a small territorial portion, these environments are important centers of multi-taxonomic biodiversity.
Keywords: Moist forests, Capões, Species richness, Conservation Units, Preservation.
Artigos
Flora arbórea de capões na Reserva da Biosfera da Serra do Espinhaço
Arboreal flora of "capões" in the Espinhaço Range Biosphere Reserve
Thaís Ribeiro Costa thais.costa@ufvjm.edu.br
Cristiane Coelho de Moura
Evandro Luiz Mendonça Machado
Anne Priscila Dias Gonzaga
Recepção: 15 Junho 2021
Aprovação: 06 Julho 2021

Nos últimos milhares de anos, a expansão das florestas em áreas de savanas e campos ocorreu em muitas partes da América do Sul, um fenômeno que tem sido geralmente atribuído às flutuações climáticas (Bueno et al. 2017; Arruda et al., 2018). No entanto, mudanças climáticas não são a única influência importante no limite destes ecótones. Outros processos, como a dispersão de espécies, características dos solos e distúrbios antrópicos também podem influenciar a distribuição da vegetação (Souza et al. 2020). Na Reserva da Biosfera da Serra do Espinhaço (RBSE), sudeste do Brasil, é possível verificar um complexo mosaico fitofisionômico decorrente de elevada heterogeneidade ambiental (i.e. variações climáticas, hidrológicas, edáficas e altitudinais) (Braga et al. 2017)
Na RBSE um sistema peculiar de floresta-campo merece destaque. Trata-se de “ilhas” de florestas estacionais naturais conhecidas como Matas de Galerias ou “Capões” (Coelho et al., 2018; Gonçalves et al., 2020). Constituem-se importantes centros de biodiversidade multitaxonômicos, apesar de ocuparem pequena extensão territorial (Fonseca et al., 2018), além de grande relevância na provisão de serviços ambientais e ecossistêmicos em múltiplas formas (Oliveira et al., 2015), especialmente no fornecimento de água (Mendonça-Galvão et al., 2011). Em todas as estações do ano, possuem, na maioria das amostragens, espécies típicas de áreas úmidas e alagadas (Silva et al., 2016).
Estes capões, comuns nas zonas de transição entre os biomas Mata Atlântica, Cerrado, Pantanal e Caatinga (Ribeiro e Walter, 2008), se enquadram como florestas estacionais do Domínio Atlântico e ocorrem associadas a uma transição brusca para áreas campestres da savana (Silveira et al., 2016), e seus limites são controlados pelo gradiente de fertilidade do solo e condições microclimáticas específicas (Coelho et al., 2018). Constituemformações florestais naturais que ocorrem em regiões deprimidas da paisagem e sob afloramento do lençol freático, que favorecem sua condição de alagamento mesmo em períodos de seca (Ribeiro e Walter, 2008; Fontes e Walter, 2011; Fontes et al., 2015).
A despeito de sua importância, estes ambientes constituem uma das formações mais ameaçadas da RBSE (Silva et al., 2016), seja pela forte atuação antrópica (Queiroz et al., 2018) e pela alta sensibilidade ao fogo (Alvarado et al., 2017), como pelo isolamento físico natural. Além disto, sua vegetação é composta por poucas espécies conspícuas, que necessitam de uma série de adaptações e atributos que permitam seu estabelecimento (Coelho et al., 2018). Assim, torna-se urgente a necessidade de preservar as mesmas, sendo apontadas como áreas prioritárias para conservação (Fonseca, 2020).
No entanto, ainda há muita negligência sobre esta fitocenose, sendo necessário pesquisas pontuais para enaltecer sua importância. Assim, o objetivo deste estudo foi realizar a caracterização florística de capões de mata (ilhas florestais) localizados em Unidades de Conservação da Reserva da Biosfera da Serra do Espinhaço. Esperamos que este artigo possa contribuir com o conhecimento da biodiversidade valiosa destes ambientes, servindo de subsídio para o estabelecimento de políticas públicas voltadas à sua conservação.
O estudo foi realizado no Parque Estadual do Biribiri (PEBi) e Parque Nacional das Sempre Vivas (PNSV), localizados na porção da Serra do Espinhaço Meridional (SdEM), na região sudeste do município de Diamantina, Minas Gerais. O clima da região é tipicamente subtropical de altitude, Cwb pela classificação de Koppen, caracterizado por verões brandos e úmidos e invernos mais frescos e secos (Sá Júnior et al., 2012).
Os quatro capões selecionados no PEB encontram-se entre as coordenadas 18°11’50,3”S 43°34’39,21”W e 18°11’20,37”S 43°34’34,6” W . Já os dezoito capões do PNSV estão localizadas na porção sul e norte, entre as coordenadas 17°55’40,7”S 43°49’48,8” W e 17°35'43.26"S 43°47'3.71"W (Figura 1).
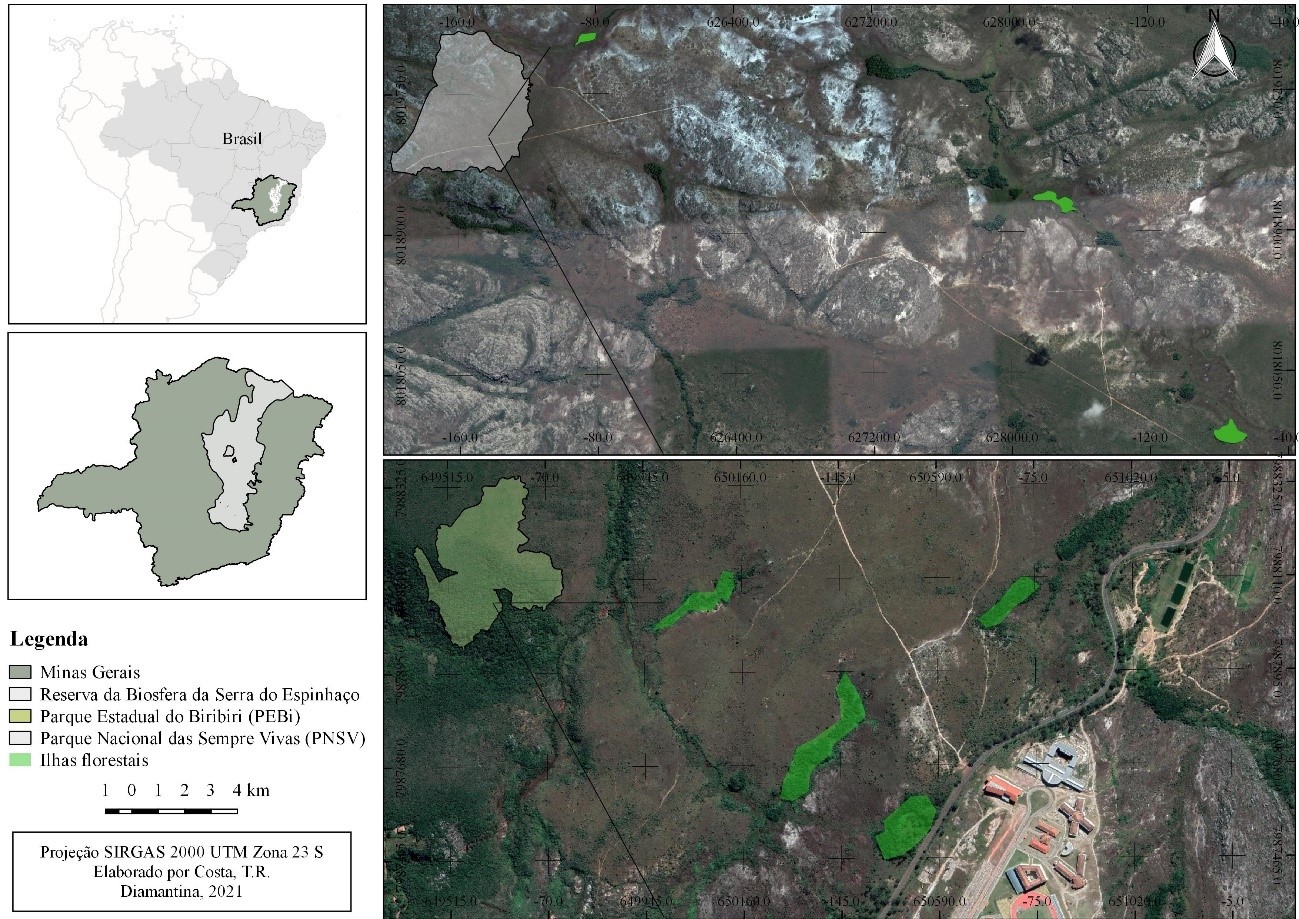
Todas as áreas amostradas estão distribuídas em altitude variando entre 1200 a 1380 m, distantes entre si, em no mínimo 200 m (Figura 1). Por estarem próximos ao topo das montanhas, estes ambientes estão dentro da zona de influência da Massa Atlântica Tropical oriunda da encosta leste (Domínio da Mata Atlântica) (Coelho et al., 2016). Ocorrem ao longo das nascentes dos cursos d’água e estão estabelecidas em manchas de Organossolos hidromórficos (Varajão et al., 2020), formando uma transição com as fitofisionomias secas adjacentes (Cerrado sentido restrito, campos rupestres e úmidos (i.e. “turfeiras”)). Assim, este sistema floresta-campo fornece importante serviço ecológico de proteção de regiões de nascentes e cabeceiras de afluentes de importantes bacias hidrográficas brasileiras (Coelho et al., 2018).
Os capões apresentam tamanho variável entre 0,02 e 3,0 ha, e se encontram em diferentes status de conservação, devido especialmente aos impactos causados pelo fogo (Figura 2). Cabe aqui a ressalva de que a prática da queimada ainda é muito comum no interior da área de proteção integral, para renovação de pastagens e coleta de sempre-vivas (Eriocaulaceae) (Sano e Giullieti, 2012). Sendo registrados no período de 2000 a 2014 aproximadamente 964 focos de incêndios dentro do território do PNSV e em suas proximidades (Soares, 2016).

Os capões foram inventariadas no mínimo 5 % de sua área total, por meio do sistema de amostragem aleatório. As parcelas permanentes tiveram dimensões de 10 x 10 m (100 m²), conforme sugerido pelo “Manual para o Monitoramento de Parcelas Permanentes nos Biomas Cerrado e Pantanal” (Felfilli et al., 2005). Assim, alocamos 72 parcelas nas ilhas florestais do PNSV, totalizando 0,72 ha de área amostral. Já no PEBi, o levantamento foi realizado em 12 parcelas (0,12 ha).
Foram amostrados todos os indivíduos com diâmetro à altura do peito maior ou igual a 3,0 cm (DAP ≥ 3,0 cm) obtidas a partir da circunferência à altura do peito (CAP). A circunferência foi medida com auxílio de fita métrica graduada em centímetros e a altura total com vara graduada em metros (projeção vertical do solo ao topo da copa). Não foram incluídas lianas e indivíduos mortos.
Foi realizada coleta do material botânico e posteriormente depositado no Herbário Dendrológico Jeanini Felfilli (HDJF), pertencente à Universidade Federal dos Vales do Jequitinhonha e Mucuri (UFVJM). A identificação das espécies, sempre que possível, foi realizada em campo, e também por meio de comparações em herbários, consulta a especialistas e bibliografia especializada.
As espécies encontradas nas ilhas florestais foram listadas em ordem de família e gênero. Para a classificação das famílias foi adotado o Angiosperm Phylogeny Group(APG) IV (2016) e os nomes de táxons foram atualizados usando o “International Plant Names Index-2012” (IPNI, 2012).
Foram realizadas consultas no Livro Vermelho da Flora do Brasil (Martinelli e Moraes, 2013) para averiguar quais espécies amostradas na área são consideradas ameaçadas de extinção. As categorias de ameaça foram definidas pela União Mundial para a Natureza (IUCN) e distribuem-se em: criticamente em perigo, em perigo, vulnerável, quase ameaçada, menos preocupante, dados insuficientes, extinta e extinta na natureza.
Para o conjunto total de capões, foram amostrados 1692 indivíduos totais, pertencentes a 40 famílias botânicas e 112 espécies (Tabela 1).
As famílias com maior riqueza foram Myrtaceae (12), Fabaceae (10), Lauraceae (10), Rubiaceae (8) e Melastomataceae (6). A maioria das espécies identificadas neste estudo, ocorreram em outros estudos de matas de galeria inundáveis ( Silva, 2012; Gaya, 2014; Costa, 2017; Moura et al. 2021), e estão geralmente associados a solos com saturação hídrica. Entre os trabalhos citados, a família Fabaceae é uma exceção, pois possui pouca importância em estudos semelhantes, uma vez que, as espécies desta família não se estabelecem plenamente em matas de galeria inundáveis (Costa, 2017), sendo a mesma, considerada a família mais representativa em termos de número de indivíduos e de espécies em matas de galeria não inundáveis (Silveira, 2020)
Apesar das matas serem caracterizadas como inundáveis, algumas parcelas se encontravam mais drenadas em relação à maioria amostrada, e estas espécies ocorreram nestes sítios, especificamente. A ocorrência de espécies desta família botânica no presente estudo pode ter sido favorecida nestas parcelas em questão, próximas às bordas das matas, uma vez que, além de serem espécies generalistas de habitat, são indicadoras de locais mais abertos (Gonçalves et al., 2016). Estes resultados evidenciam que a maior heterogeneidade ambiental favorece a ocorrência de diferentes espécies, com requerimentos ambientais específicos, aumentando a riqueza em áreas com maior diversidade de ambientes (Silveira, 2020).
As espécies que mais se destacaram, em relação ao número de indivíduos foram: Richeria grandis ocupando o primeiro lugar no rank com 252 indivíduos, Xylopia emarginata (182 ind.), Protium spruceanum (149 ind.), Swartia multijuga (116 ind.) e Trembleyaa parviflora (102 ind.) (Tabela 1).
Moura et al. (2021) e Costa (2017) em seus estudos, apontaram a R. grandis como a espécie de maior valor de importância, corroborando para o resultado deste estudo. R. grandis apesar de ter uma especificidade ambiental única, apresenta uma distribuição geográfica ampla (Rabinowitz, 1981). Deste modo, é considerada uma das espécies mais representativas de ambientes ripários, caracterizados por solos hidromórficos e distróficos (Costa, 2017; Huber e Rull, 2019), uma vez que possui adaptações morfofisiológicas para suportar as condições físicas e a baixa oxigenação do solo nestes locais (Bambi et al., 2016), e é responsável por contribuir com mais de 45% da biomassa da serapilheira, determinante para a estabilidade destes ecossistemas (Rezende et al., 2017; Calderón et al. 2019).

As espécies Ferdinandusa speciosa, Geonoma schotiana, Cecropia pachystachya, Hedyosmum brasiliensis, além das supracitadas, são comumente abordadas na maioria das pesquisas desenvolvidas em ecossistemas de florestas ripárias (Gonzaga et al, 2019), onde figuram entre as mais abundantes. Dentre as espécies amostradas, algumas podem ser consideradas generalistas por compartilhar áreas de florestas e cerrado (Mendonça, 2008) como Copaifera langsdorffi, Maprounea guianensis, Trembleya parviflora, Tabebuia ocharea, Eremanthus incanus, dentre outras. Isto ressalta a influência das formações vegetais adjacentes na composição e diversidade destas florestas. Este resultado sugere que estes ambientes podem atuar como corredores ecológicos entre diferentes fitocenoses, e, portanto, pode constituir importante área de refúgio frente às futuras mudanças climáticas.
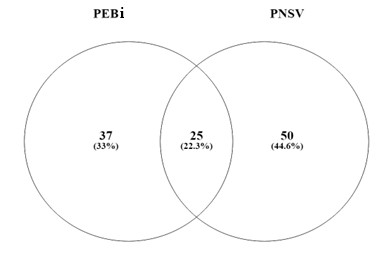
Das espécies amostradas, 50 ocorreram exclusivamente nos capões do PNSV, enquanto 37 espécies ocorreram apenas no PEBi. As duas áreas compartilharam apenas 25 espécies (22,3%) em comum (Figura 3). Este resultado sugere elevada diversidade beta entre as áreas, o que ressalta a importância singularidade e contribuição de cada Unidade de Conservação para a biodiversidade regional. Logo, estas áreas apresentam valor inestimável para manutenção dos processos de evolução (como especiação, fluxo gênico e adaptação) em zonas ecotonais (Souza, 2020).
O conhecimento da flora destes capõesnaturalmente isolados permite a extrapolação dessas informações para um plano geográfico, sendo uma ferramenta de grande valor prático para apoio à tomada de decisão. Com base nisso, evidenciamos a importância das seguintes medidas: i) manutenção e ampliação das Unidades de Conservação em locais com elevada diversidade ii) garantia da conectividade da paisagem por meio de corredores ecológicos (Morandi et al. 2020), visando favorecer o fluxo das espécies para diferentes fragmentos de floresta.
De acordo com Martins (1991), as espécies consideradas raras são aquelas que apresentam apenas um indivíduo na área amostrada. Com isso, as áreas deste estudo apresentaram 31 espécies raras, que representam 27,67 % das espécies amostradas e 1,18% dos indivíduos (Tabela 1).
Espécies que apresentam baixa densidade podem indicar duas situações distintas: i) que não tiveram muito sucesso na capacidade de colonização na comunidade, ou ii) pode tratar de um efeito da competição interespecífica dessas com as espécies já estabelecidas e melhor adaptadas às condições de alagamento do solo da área de estudo. Este padrão de ocorrência com muitas espécies representadas por poucos indivíduos apresenta fragilidade adicional dos ecossistemas fragmentados, pois a perda de alguns indivíduos pode ocasionar a perda de diversidade de espécies (Fagundes et al., 2019).
Dentre as espécies amostradas, quinze estão inseridas no Livro Vermelho da Flora do Brasil (2013), sendo estas: Euterpe edulis e Machaerium villosum categorizadas como vulneráveis e Nectandra warmingi, considerada como quase ameaçada. As espécies Andira fraxinifolia, Cletra scraba, Copaifera langsdorfii, Dendropanax cuneatum, Erythroxylum pelletelianum, Geonoma schotiana, Inga marginata, Maytenus robusta, Ocotea puberula e Mgnolia ovata estão inclusas na categoria de menos preocupante e Ocotea acyphila e Protium heptaphylum são consideradas espécies com deficiência de dados.
A composição de espécies categorizadas como ameaçadas de extinção e o elevado número de espécies consideradas raras acentuam, portanto, a importância da flora arbórea dos capões no Espinhaço.
Nossos resultados demonstraram que capões naturalmente fragmentados da Reserva da Biosfera da Serra do Espinhaço são ambientes que contribuem significativamente para a biodiversidade regional, uma vez que constituem importantes centros multitaxonômicos da flora. Adicionalmente, a identificação de espécies raras e ameaçadas de extinção indicam a urgência de proteção efetiva destas florestas. Esperamos que este trabalho possa nortear pesquisas futuras que garantam a conectividade destas florestas, como métricas de ecologia da paisagem para implementação de corredores ecológicos em Unidades de Conservação.
O presente trabalho foi realizado com apoio da Coordenação de Aperfeiçoamento de Pessoal de Nível Superior -Brasil (CAPES) - Código de Financiamento 001.


