Abstract:
Two experiments
were carried out aiming to study the value of whole oats (wo) as a potential source of physically
effective fiber (pendf) in rations
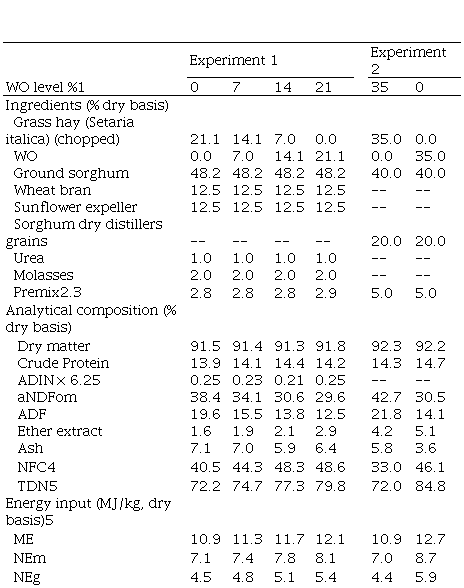
for calves. In experiment 1, 24 Hereford calves (177 ± 18 kg) were randomly
assigned to eight groups and these into one of four experimental rations (79%
concentrate), varying in the replacement level of grass hay (gh) for wo: 0:21, 7:14, 14:7, 21:0% dry matter
(dm) basis wo:gh,
respectively. Increasing wo level
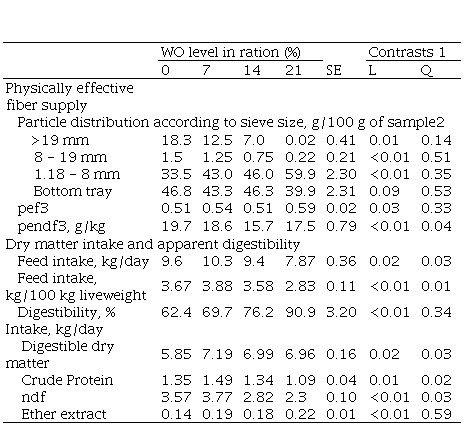
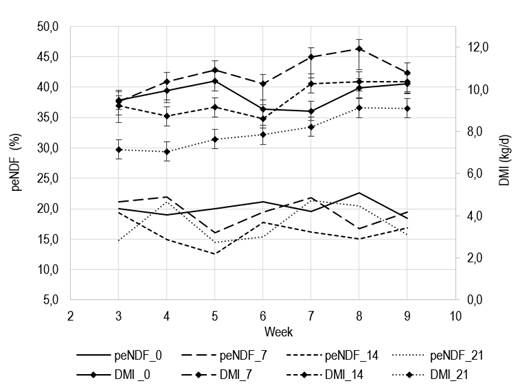
resulted in a quadratic response for pendf (P= 0.04) with
values above 15% of dm. Intake
increased quadratically (P= 0.03) and dm
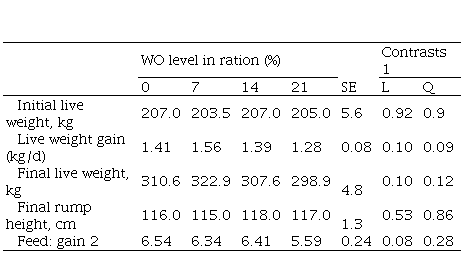
digestibility increased linearly (P= 0.03). As a result, feed conversion
efficiency tended to improve (P= 0.08). In Experiment 2, four rumen cannulated
steers were assigned in a crossover design to one of two experimental rations
(65% concentrate) varying in pendf fiber source:
35% gh
or 35% wo, aiming to study the
fermentative pattern. Replacing gh for wo
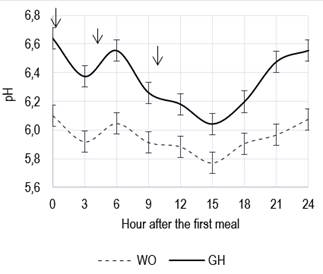
reduced mean daily rumen pH (5.9 vs. 6.3, P < 0.01), varying between 6.1 and
5.8, and always below pH values in gh. No differences were observed for the potentially
degradable dm fraction, but
fermentation rate was higher for wo
(35.0 vs. 9.0 %/ h; P= 0.02), hence increasing effective degradability (58.3
vs. 65.4%; P= 0.048). Results from both experiments suggest it is viable to use
wo as a pendf source for
concentrated rations.
Keywords: cattle, growth, effective fiber, digestion, feed conversion.
Resumen:
Se realizaron dos experimentos con el objetivo de estudiar el valor del grano entero de avena (gea) como fuente de fibra físicamente efectiva (pefdn) en raciones para terneros. En el experimento 1, 24 terneros Hereford (177±18 kg) fueron sorteados a ocho grupos y estos a una de cuatro raciones (79 % concentrado), definidas por niveles crecientes de sustitución de heno de moha (hm) por gea: 0:21, 7:14, 14:7, 21:0 % base materia seca (ms) gea:hm, respectivamente. Niveles crecientes de gea resultaron en una respuesta cuadrática de pefdn (P= 0,04) ubicándose por encima del 15 %. El consumo aumentó en forma cuadrática (P= 0,03) y la digestibilidad linealmente (P= 0,03). Como resultado, a mayor gea la eficiencia de conversión tendió a mejorar (P= 0,08). En el Experimento 2, cuatro novillos provistos de cánula ruminal se asignaron en un diseño crossover a dos raciones (65 % concentrado), difiriendo en la fuente de pefdn: 35 % hm o 35 % gea, con el objetivo de estudiar sus características fermentativas. La sustitución de hm por gea redujo el pH medio del rumen (5,9 vs. 6,3, P <0,01) variando entre 6,1 y 5,8, permaneciendo las 24 h por debajo de los valores registrados para hm. No se afectó la fracción de la ms potencialmente degradable en rumen, pero aumentó la tasa de fermentación para gea (35,0 vs. 9,0 %/h; P= 0,02), y la degradabilidad efectiva (58,3 vs. 65,4 %; P= 0,048). Los resultados de ambos experimentos sugieren que es viable el uso del gea como fuente de pefdn en raciones concentradas.
Palabras clave: vacuno, crecimiento, fibra efectiva, digestión, eficiencia de conversión.
Resumo:
Dois experimentos foram conduzidos como o objetivo de estudar o valor do grão inteiro de aveia (GIA) como fonte de fibra fisicamente efetiva (peFDN) em rações para bezerros. No experimento 1, 24 bezerros Hereford (177 ± 18 kg) foram sorteados em oito grupos e estes assignado ao acaso a quatro rações experimentais (79% de concentrado), definidas pelo aumento dos níveis de substituição de feno de moha (FM) pelo GIA: 0 : 21, 7:14, 14: 7, 21: 0% da matéria seca (MS) GIA: FM, respectivamente. O aumento do GIA resultou numa resposta quadrática de peFDN (P= 0,04) atingindo mais de 15%. O consumo aumentou quadraticamente (P= 0,03) e a digestibilidade linearmente (P= 0,03). Como resultado, quanto maior o GIA, a conversão alimentar tendeu a melhorar (P= 0,08). No experimento 2, quatro novilhos portadores de cânula ruminal foram designados num delineamento crossover a duas rações (65% concentrado) diferindo na fonte de peFDN: 35% FM ou 35% GIA, a fim de estudar as características fermentativas. A substituição de FM por GIA reduziu o pH médio do rúmen (5,9 vs. 6,3, P<0,01), variando entre 6,1 e 5,8, permanecendo 24 horas abaixo dos valores de FM. A fração da MS potencialmente degradável no rúmen não foi afetada, mas o GIA aumentou a taxa de fermentação (35,0 vs 9,0% / h; P= 0,02), e a degradabilidade efetiva (58,3 vs 65,4%; P= 0,048). Os resultados de ambos os experimentos sugerem que é viável o uso de GIA como fonte de peFDN em rações concentradas.
Palavras-chave: bovinos, crescimento, fibra eficaz, digestão, eficiência de conversão.
References:
1. Parra V, Rifle SL, Elizalde JC. Estrategias de inclusión del corral en los sistemas ganaderos de la Argentina. Balcarce: the authors; 2006. 180p.
2. Simeone A, Beretta V, Elizalde JC. Encierre de terneros o sistema ADT. In: Simeone A, Beretta V, editors. Jornada Anual de la Unidad de Producción Intensiva de Carne: una década de investigación para una ganadería más eficiente. Paysandú: Facultad de Agronomía; 2008. p. 38-41.
3. Mertens DR. Measuring fiber and its effectiveness in ruminant diets. In: Proceedings of Plains Nutrition Conference; 2002 Apr 25-26; San Antonio, Texas [Internet]. Texas: Texas A&M Research and Extension Center; 2002 [cited 2020 Sep 25]. p. 40-66. Available from: https://bit.ly/368PbWi.
4. Fox DG, Tedeschi LO. Application of physically effective fiber in diets for feedlot cattle. In: Proceedings of Plains Nutrition Conference; 2002 Apr 25-26; San Antonio, Texas [Internet]. Texas: Texas A&M Research and Extension Center; 2002 [cited 2020 Sep 25]. p. 67-81. Available from: https://bit.ly/2EBQ7qK.
5. Beauchemin KA, Yang WZ. Effects of Physically Effective Fiber on Intake, Chewing Activity, and Ruminal Acidosis for Dairy Cows Fed Diets Based on Corn Silage. J Dairy Sci. 2005;88:2117-29.
6. Mertens DR. Creating a system for meeting the fiber requirements of dairy cows. J Dairy Sci. 1997;80:1463-81.
7. Beretta V, Simeone A, Elizalde JC, Franco J, Bentancur O, Ferrés A, Aycaguer S, Iriñiz J, Martínez V. Alternative fiber sources for steers and calves fed high grain feedlot diets. Anim Prod Sci. 2010;50:410-3.
8. Simeone A, Beretta V. Evaluation of a self-feeding system with a total mixed ration without long fiber to lot-fed beef calves. Agrociencia Uruguay. 2018;22(2):1-9.
9. Beretta V, Simeone A, Franco J, Casaretto F, Mondelli S, Valdez G. Using self-feeders with high concentrate diets for finishing beef cattle. In: Book of Abstracts of 69th Meeting of the European Federation of Animal Science. Wageningen: Wageningen Academic Publishers; 2018. p. 486.
10. Herrera-Saldana RE, Huber JT, Poore MH. Dry Matter, Crude Protein, and Starch Degradability of Five Cereal Grains. J Dairy Sci. 1990;73(9):2386-93.
11. Pordomingo AJ, Jonas O, Adra M, Juan NA, Azcárate MP. Evaluación de dietas basadas en grano entero, sin fibra larga, para engorde de bovinos a corral. Rev investig agropecu. 2002;31(1):1-23.
12. Morgan CA, Campling RC. Digestibility of whole barley and oat grains by cattle of different ages. Anim Sci. 1978;27(3):323-9.
13. National Academies of Sciences, Engineering, and Medicine. Nutrient Requirements of Beef Cattle. 8th rev ed. Washington: The National Academies Press; 2016. 494p.
14. Guidelines for uniform beef improvement programs [Internet]. 9th ed. Prairie (MS): Beef Improvement Federation; 2016 [cited 2020 Sep 23]. 185p. Available from: https://bit.ly/3hTCall.
15. Heinrichs J, Kononoff P. Evaluating particle size of forages and TMRs using the New Penn State Forage Particle Separator [Internet]. Pennsylvania: Pennsylvania State University; 2002 [cited 2020 Sep 25]. 14p. (DAS; 02- 42). Available from: https://bit.ly/340aMxm.
16. Van Keulen J, Young BA. Evaluation of acid-insoluble ash as a natural marker in ruminant digestibility studies. J Anim Sci. 1977;44:282-7.
17. Latimer GW, editor. Official methods of analysis. 19th ed. Gaithersburg (MD): AOAC; 2012. 2v.
18. Van Soest PJ, Robertson JB, Lewis BA. Methods for dietary fiber, neutral detergent fiber and nonstarch polysaccharides in relation to animal nutrition. J Dairy Sci. 1991;74:3583-97.
19. Tejada de Hernández I. Manual de laboratorio para análisis de ingredientes utilizados en la alimentación animal. México: Instituto Nacional de Investigaciones Pecuarias; 1983. 387p.
20. Ørskov ER, Hovell FD, Mould F. The use of the nylon bag technique for evaluation of feedstuffs. Trop Anim Prod. 1980;5(3):195-213.
21. Trujillo AI, Bruni M, Chilibroste P. Nutrient content and nutrient availability of sorghum wet distiller’s grain in comparison with the parental grain for ruminants. J Sci Food Agric [Internet]. 2016 [cited 2020 Sep 25];97(8):2353-7. Available from: https://bit.ly/342kj75.
22. Ørskov ER, McDonald I. The estimation of protein degradability in the rumen from incubation measurements weighted according to rate of passage. J Agric Sci. 1979;92:499-503.
23. AFRC. Energy and Protein requirements of ruminants. Cambridge: CAB International; 1993. 159p.
24. Zinn RA. Influence of Processing on the Feeding Value of Oats for Feedlot Cattle. J Anim Sci. 1993;71:2303-9.
25. Arelovich HM, Abney CS, Vizcarra JA, Galyean ML. Effects of dietary neutral detergent fiber on intakes of dry matter and net energy by dairy and beef cattle: analysis of published data. Prof Anim Sci. 2008;24:375-83.
26. McAllister TA, Gibb DJ, Beauchemin KA, Wang Y. Starch type, structure and ruminal digestion. In: Cattle Grain Processing Symposium; 2006 Nov 15-17; Tulsa, Oklahoma, United States [Internet]. Stillwater: University of Oklahoma; 2007 [cited 2020 Sep 25]. p. 30-41. Avalilable from: https://bit.ly/33XITpI.
27. Thonney ML, Hogue DE. Formulation of ruminant diets using by-product ingredients on the basis of fermentable NDF and nonstructural carbohydrate. In: Cattle Grain Processing Symposium; 2006 Nov 15-17; Tulsa, Oklahoma, United States [Internet]. Stillwater: University of Oklahoma; 2007 [cited 2020 Sep 25]. p. 205-13. Avalilable from: https://bit.ly/33XITpI.
28. Owens FN, Secrist DS, Hill WJ, Gill DR. The Effect of Grain Source and Grain Processing on Performance of Feedlot Cattle: a Review. J Anim Sci. 1997;75:868-79.
29. Owens FN, Soderlund S. Ruminal and post ruminal starch digestion by cattle. In: Cattle Grain Processing Symposium; 2006 Nov 15-17; Tulsa, Oklahoma, United States [Internet]. Stillwater: University of Oklahoma; 2007 [cited 2020 Sep 25]. p. 116-28. Avalilable from: https://bit.ly/33XITpI.
30. Beauchemin KA, McAllister TA, Dong Y, Farr BI, Cheng KJ. Effects of mastication on digestion of whole cereal grains by cattle. J Anim Sci. 1994;72:236-46.
31. Huntington GB, Harmon DL, Richards CJ. Sites, rates, and limits of starch digestion and glucose metabolism in growing cattle. J Anim Sci. 2006;84(E. Suppl.):E14-24
32. Owens FN, Zinn RA, Kim YK. Limits to starch digestion in the ruminant small intestine. J Anim Sci. 1986;63:1634-48
33. Allen MS. Drives and limits to feed intake in ruminants. Anim Prod Sci. 2014;54:1513-24.
34. Miner JL. Recent Advances in the central control of intake in ruminants. J Anim Sci. 1992;70:1283-9.
35. Vasconcelos JT, Galyean ML. Technical Note: do dietary net energy values calculated from performance data offer increased sensitivity for detecting treatment differences? J Anim Sci. 2008;86:2756–60.
36. Ørskov ER, Flatt WP, Moe PW. Fermentation balance approach to estimate extent of fermentation and efficiency of volatile fatty acid formation in ruminants. J Dairy Sci. 1968;51:1429-35.
37. González LA, Manteca X, Calsamiglia S, Schwartzkopf-Genswein KS, Ferret A. Ruminal acidosis in feedlot cattle: interplay between feed ingredients, rumen function and feeding behavior. Anim Feed Sci Tech. 2012;172:66-79.
38. Stock R, Britton R. G91-1047 Acidosis. In: Historical Materials from University of Nebraska-Lincoln Extension [Internet]. 1991 [cited 2020 Sep 25]. 8p. Available from: https://bit.ly/2RWeDG7.
39. Pacheco RDL, Cruz GD. Acidosis in cattle. In: Puniya AK, Singh R, Kamra DN, editors. Rumen Microbiology: from evolution to revolution. New Delhi: Springer; 2015. p. 315-27.
40. National Research Council. Nutrient requirements of beef cattle. 7th ed. Washington: National Academic Press; 1996. 248p.
41. Schwartzkopf-Genswein KS, Beauchemin KA, Gibb DJ, Crews DH, Hickman DD, Streeter M, McAllister TA. Effect of bunk management on feeding behavior, ruminal acidosis and performance of feedlot cattle: a review. J Anim Sci. 2003;81(E. Suppl. 2):E149-58.
42. Simeone A, Beretta V, Elizalde JC, Franco J, Viera G. Effect of removing long fibre from beef cattle feedlot diets. In: Book of abstracts for the 10th Word Conference on animal Production. Netherlands: Wageningen Academic Publishers; 2008. p. 130.
Notes
Author notes
virginia.beretta@gmail.com
 Beretta, V. virginia.beretta@gmail.com
Beretta, V. virginia.beretta@gmail.com