Resumen: El género neotropical Varronia P. Browne es uno de los más problemáticos, taxonómicamente, entre las Boraginales. Esto puede deberse a la frecuente superposición de los límites entre la variación intra e interespecífica de algunos caracteres. La finalidad de este trabajo fue explorar el potencial de los caracteres morfoanatómicos foliares para la delimitación de las especies que se encuentran dentro del complejo Varronia bullata en Panamá: Varronia inermis (Mill.) Borhidi, Varronia bullata L. y Varronia linnaei (Stearn) J.S. Mill. Para esto, se colectaron hojas maduras en diferentes poblaciones de cada especie. Se realizaron cortes transversales y se obtuvieron secciones superficiales de la lámina foliar, los cuales fueron utilizados para cuantificar y cualificar caracteres morfoanatómicos previamente seleccionados. La información cuantitativa fue sometida a un análisis canónico discriminante. Los resultados del análisis canónico discriminante mostraron que la densidad de los tricomas simples en la epidermis abaxial y la longitud de los tricomas simples en la epidermis adaxial, son los caracteres que presentan mayor influencia en la separación de las especies; por otro lado, a manera descriptiva se refleja que la presencia de sales cristalinas, el tipo de tricomas y la distribución de los estomas también contribuyen en la separación taxonómica de las especies.
Palabras clave: Boraginales, superficie foliar, tricomas .
Abstract: The neotropical genus Varronia P. Browne is one of the most problematic, taxonomically, among the Boraginales. This may be due to the frequency of overlapping the boundaries between intra and interspecific variation of some characters. The purpose of this project was to explore the potential of foliar morpho-anatomic characters for the delimitation of the species found within the Varronia bullata complex in Panama: Varronia inermis (Mill.) Borhidi, Varronia bullata L. and Varronia linnaei (Stearn) J.S. Windmill. For this purpose, mature leaves were collected in different populations for each species. Cross sections were made, and superficial sections of the leaf lamina were obtained, which were used to quantify and qualify previously selected morphoanatomic characters. The quantitative information was subjected to a discriminant canonical analysis. The results of the discriminating canonical analysis that detect the density of simple trichomes in the abaxial epidermis and the length of simple trichomes in the adaxial epidermis, are the characters that have the greatest influence on species separation. On the other hand, a descriptive way is reflected that the presence of crystalline sales, the type of trichomes and the distribution of stomata also contribute to the taxonomic separation of species.
Keywords: Boraginales, leaf surface, trichomes .
Artículos
MORFOANATOMÍA FOLIAR COMO EVIDENCIA TAXONÓMICA EN LA DELIMITACIÓN DE LAS ESPECIES QUE FORMAN EL COMPLEJO Varronia bullata (CORDIACEAE)
LEAF MORPHOANATOMY AS TAXONOMIC EVIDENCE IN THE DELIMITATION OF THE SPECIES INCLUDED IN THE Varronia bullata COMPLEX
Recepción: 27 Julio 2020
Aprobación: 06 Noviembre 2020
El género Varronia P. Browne perteneciente a la familia Cordiaceae, se distribuye desde las regiones cálidas de Arizona hasta Argentina (Miller y Gottschling, 2007). Según Estrada (1995), se reconocen tres áreas de gran diversidad para Varronia, la primera de ellas se extiende por Brasil, la segunda está en México y la tercera se ubica a lo largo de Los Andes en Chile. El género se caracteriza por estar constituido por arbustos con hojas aserradas y venación craspedódroma, inflorescencias en cimas bifurcadas, sindesmios espiciformes o capituliformes, estigmas dos veces bífido, polen triporado y reticulado, frutos carnosos de color rojo y mesocarpo mucilaginoso o jugoso (Campos y Chiang, 2012).
Si bien, el género Varronia fue propuesto por Patrick Browne (1756), algunos autores posteriores lo consideraron como parte de Cordia L. (Johnston, 1930; Miller, 1988; Estrada, 1995), debido a caracteres como el número de estigmas (4) y el tipo de fruto (drupáceos). Borhidi et al. (1988) analizaron caracteres como la morfología del polen, la anatomía floral (especialmente la apertura del cáliz) y la morfología de los lóbulos del cáliz; y concluyeron que se podían diferenciar tres taxones: Cordia L., Varronia P. Browne y Gerascanthus P. Browne. En base a estos resultados, Nowicke y Miller (1990) reconocieron que Varronia debía ser considerada genéricamente distinta a Cordia. Hoy día, el género Varronia se considera un grupo monofilético (Gottschling et al., 2005; Miller y Gottschling, 2007), lo cual es sustentado por caracteres polínicos (Borhidi et al., 1988).
En 1969, Nowicke realizó la primera revisión de la familia Boraginaceae (incluido el género Varronia) para la Flora de Panamá. Luego, Gentry y Janos (1974) publicaron una lista preliminar para América Central, donde reportaron 17 especies de Cordia (incluyendo Varronia) para Panamá. Recientemente, en el tratamiento para la Flora Mesoamericana, Miller (2011) reconoce la familia Cordiaceae, y en ella, incluye los géneros Cordia y Varronia. Este autor registra siete especies de Varronia para Panamá: Varronia bullata L., Varronia curassavica Jacq., Varronia inermis (Mill.) Borhidi, Varronia linnaei (Stearn) J.S. Mill., Varronia globosa Jacq., Varronia spinescens (L.) Borhidi y Varronia dichotoma Ruiz y Pav.
De las especies del género Varronia presentes en Panamá, aún existen problemas de delimitación entre V. bullata L., V. inermis (Mill.) Borhidi y V. linnaei (Stearn) J.S. Mill. Para este estudio, las especies mencionadas fueron consideradas como el complejo Varronia bullata. El carácter más relevante que unifica al complejo es la presencia de inflorescencias de tipo cima capituliforme.
Para separar entre los taxones V. linnaei, V. inermis y V. bullata, la Flora Mesoamericana (Miller, 2011) utiliza la longitud del pedúnculo y la forma de los ápices de los lóbulos del cáliz. Sin embargo, estos caracteres son muy variables porque dependen del estado de madurez de las inflorescencias, según observaciones preliminares realizadas por los autores. Por lo tanto, utilizar los caracteres antes mencionados no es un criterio suficiente para delimitar taxonómicamente y conocer el número real de taxones que conforman este grupo.
Buscando otras alternativas para una mejor identificación, se propone la utilización de la morfoanatomía foliar como una herramienta que podría contribuir a una mejor separación de estas especies. Este carácter podría representar evidencias valiosas que permitan delimitar con más propiedad a las especies que forman el complejo Varronia bullata y eviten confusiones al momento de realizar investigaciones científicas y hacer planes de conservación para dichas especies. Por último, dada la escasez de este tipo de estudios en el grupo, es de interés desarrollar trabajos que aporten conocimientos sobre la anatomía foliar propia de los taxones involucrados.
En este sentido, la finalidad de este trabajo fue explorar el potencial de los caracteres morfoanatómicos en la delimitación de las especies que forman el complejo Varronia bullata.
Área de estudio
Para cada especie en estudio, se colectó material fresco de diferentes poblaciones. Las colectas se realizaron entre los meses de junio a diciembre del 2017. Las colectas estuvieron sujetas a la disponibilidad de las poblaciones y a la accesibilidad a los sitios. Varronia bullata se colectó en Santa Fe, Veraguas. Varronia linnaei se colectó en El Llano-Cartí, Panamá y en Santa Fe, Veraguas. Varronia inermis se colectó en los corregimientos de Limones y Alanje, y en un sitio cercano al río Guanábano (Chiriquí) (Fig. 1).
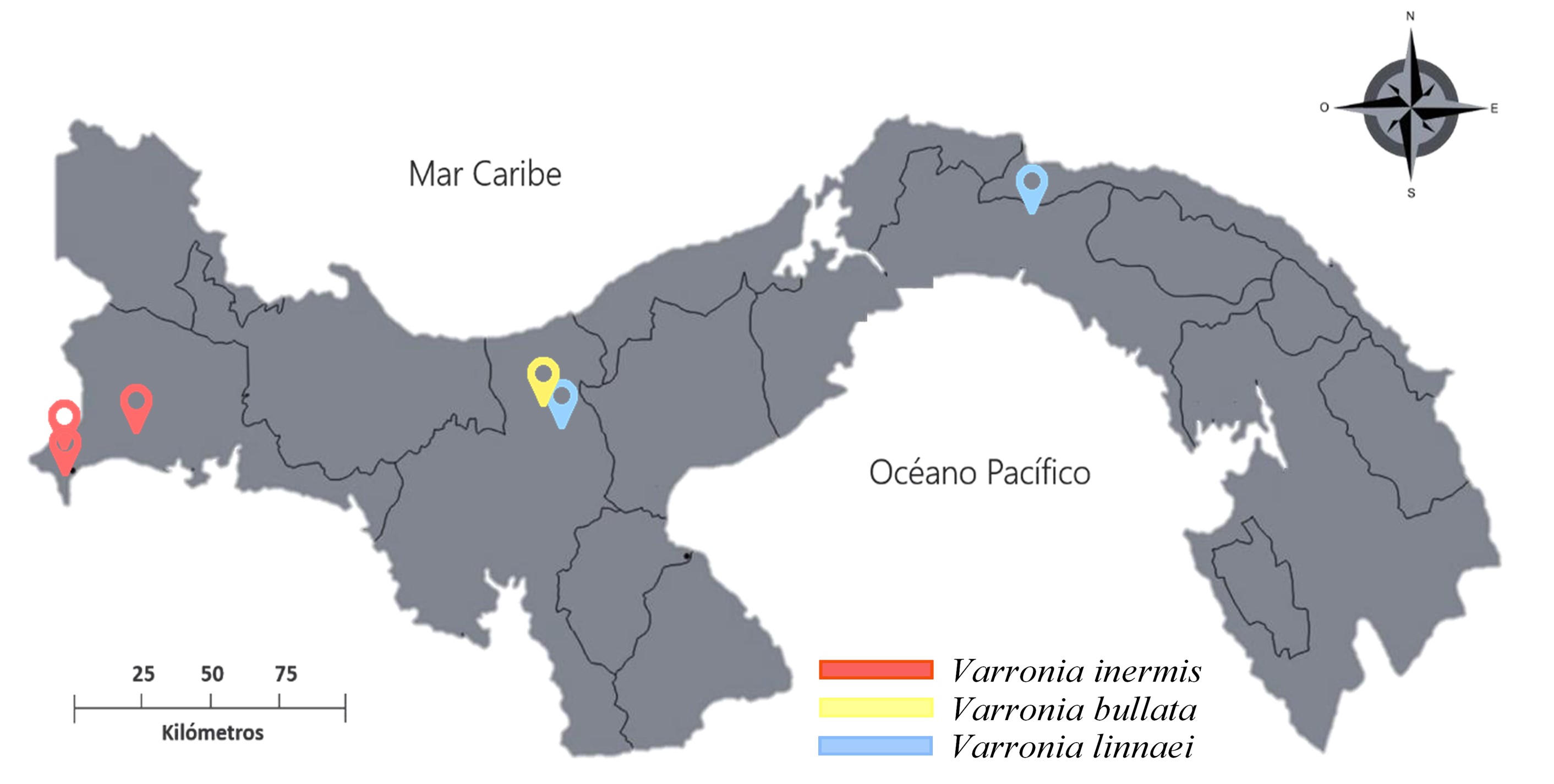
Fig. 1
Localidades de colecta de las especies estudiadas
Trabajo en campo
Para el trabajo en campo, de cada población se seleccionaron 20 individuos y por cada individuo se colectó una hoja madura de la parte superior, y otra hoja de la parte inferior de la planta. Al momento de la colecta, las hojas fueron almacenadas y fijadas en FAA (Johansen, 1940) por 24-48 horas, para luego ser procesadas en los laboratorios 7-204 y 7-39 de la Escuela de Biología, Universidad de Panamá.
Trabajo en laboratorio
El trabajo en el laboratorio se dividió en dos grandes estudios: la estructura interna y la superficie de la hoja. Para realizarlos, se obtuvieron secciones de un centímetro de ancho de la parte media de la hoja, los restos de los ápices y las bases fueron almacenados en etanol 70° para su uso en la microscopía electrónica.
Estructura interna de la hoja
Para el estudio de la estructura interna, se utilizó la metodología de Johansen (1940), modificando los tiempos de deshidratación e inclusión en parafina.
Se realizaron tres cortes transversales de 10 µm de ancho por cada sección foliar. La tinción usada consistió en azul alcián en solución acuosa 1% durante 60 minutos y safranina acuosa al 0.1% durante 90 segundos.
Las muestras fueron montadas en Bálsamo de Canadá.
Superficie foliar
Para estudiar la superficie foliar se utilizaron los métodos de diafanización y microscopía electrónica de barrido (MEB).
Diafanización
Para cuantificar los datos, se empleó el método de Stittmatter y Dizeo (1973), con modificaciones propias. Las muestras fueron teñidas con azul de toluidina en buffer de fosfato (O’Brien et al., 1964) durante 2 horas.
Al final, se separaban las epidermis del mesófilo y se colocaban sobre un portaobjeto cuidando que la superficie a observar quedara hacia arriba.
Microscopía electrónica de barrido
Las muestras fueron lavadas en etanol 50° durante una noche, luego fueron deshidratadas en cambios de etanol 70°, 80°, 90° y 100° durante una hora. Se pasaron a una solución (1:1) de Etanol-Acetato de amilo y luego a Acetato de amilo. Las muestras se secaron a punto crítico, para luego ser colocadas sobre un adhesivo de carbón en el portamuestras y pasar a ser cubiertas con una aleación de oro/paladio. Por último, se realizaron las observaciones y las microfotografías empleando un microscopio ZEISS EVO 40 VP, en el Laboratorio de Microscopía del Instituto Smithsonian de Investigaciones Tropicales.
Observaciones y mediciones
Todas las placas preparadas fueron observadas en un microscopio OLYMPUS CX21 adaptado a una cámara AmScope mu900. Los conteos y las mediciones celulares fueron realizados con el programa informático AmScope 3.7.7934, calibrado con un portaobjeto milimetrado.
Para el estudio fueron tomados en cuenta caracteres cualitativos y cuantificados. Estos últimos, fueron seleccionados de acuerdo a la importancia sistemática que poseen para la identificación en la familia Boraginaceae s.l., según Metcalfe y Chalk (1950) e incluyen modificaciones propias.
El listado de los caracteres que fueron medidos se presenta a continuación: densidad de los tricomas simples en la epidermis abaxial, densidad de los tricomas glandulares en la epidermis abaxial, densidad estomática, longitud de los tricomas simples en la epidermis abaxial, densidad de los tricomas simples en la epidermis adaxial, longitud de los tricomas simples en la epidermis adaxial, grosor de la vena media, grosor del parénquima de empalizada, grosor del parénquima esponjoso, grosor de la epidermis adaxial, grosor de la epidermis abaxial.
Análisis de datos
Con el propósito de corroborar que los individuos estuviesen correctamente asignados a las especies identificadas y, además de obtener las dos funciones discriminantes que nos permitieran determinar cuáles eran las variables con mayor influencia en la separación de las especies, se realizó un análisis discriminante clasificatorio utilizando los datos obtenidos de las mediciones.
Este análisis fue realizado con el programa estadístico Statgraphics Centurion XVII versión 17.1.12.
A continuación, se presentan los resultados obtenidos a través de las observaciones y mediciones para los caracteres morfoanatómicos de la lámina foliar de las especies de Varronia, pertenecientes al complejo Varronia bullata: V. inermis, V. linnaei y V. bullata.
Comparación de la estructura interna de la hoja entre las especies de Varronia.
Los resultados demuestran que las especies estudiadas de Varronia presentan similitudes en su estructura interna, teniendo en cuenta el número de capas celulares en la epidermis (uniestratificada), la simetría del mesófilo (dorsiventral) y la presencia de una vaina parenquimática alrededor de los haces vasculares, lo que permite sugerir cierta similitud anatómica dentro del grupo. No obstante, la presencia de sales cristalinas en la epidermis y en el mesófilo varía entre las especies.
Epidermis (en vista transversal)
Las especies estudiadas mostraron una epidermis uniestratificada, formada por células cuadrangulares a isodiamétricas, de pared tangencial externa levemente engrosada (Fig. 2D, 2E, 2F).
Mesófilo
En las especies estudiadas, el mesófilo es bifacial con simetría dorsiventral, distinguiéndose 2 tipos de parénquima, en empalizada y esponjoso. El parénquima en empalizada está constituido por un estrato de células. Las células del clorénquima, varían desde muy cortas y prácticamente isodiamétricas hasta alargadas en el sentido adaxial-abaxial de la hoja. Mientras que, el parénquima esponjoso está compuesto de dos a cuatro estratos de células de forma isodiamétrica, de tamaño variable, compactas, con paredes delgadas y grandes espacios intercelulares (Fig. 2D, 2E, 2I).
Nervadura central
Las especies estudiadas presentaron una nervadura central tipo Vitis según la clasificación de Esau (1977). La nervadura central es convexa en ambas caras, y está constituida por una epidermis uniestratificada. El haz vascular colateral cerrado se encuentra rodeado por tejido parenquimático (Fig. 2A, 2B, 2C). El arco central presenta haces continuos en V. linnaei (Fig. 2B), mientras que en V. inermis y V. bullata los haces se encuentran separados por tejido parenquimático (Fig. 2A, 2C).
Cistolitos
Todas las especies presentaron tricomas simples con bases cistolíticas. Los cistolitos se encuentran en ambas epidermis, aunque son más frecuentes en la epidermis adaxial. El tamaño de estos cistolitos varía para cada especie (Fig. 2G, 2H, 2I, 2L).
Sales cristalinas
Dentro del mesófilo y las células epidérmicas de V. bullata y en el mesófilo de V. linnaei se detectó la presencia de células con paredes delgadas de mayor tamaño que las células parenquimáticas, las cuales contenían sales cristalinas condensadas, a manera de arena cristalina (Fig. 2G, 2E). La cantidad, tamaño, forma y distribución de estas sales, varía con la población y/o ejemplar estudiado. Metcalf y Chalk (1950), en su enciclopedia “Anatomy of the Dicotyledons” realizaron una revisión con referencias dispersas sobre la anatomía de Cordia. Su breve sinopsis nos muestra la alta variabilidad que presenta la anatomía en las hojas y el tallo para diferentes especies de Cordia. Además, les llamó inmensamente la atención, la presencia de inclusiones cristalinas conspicuas en la hoja, el tallo, y las células del parénquima de la madera.
Para Gottwald (1983), el género Varronia, a diferencia de los otros géneros de Boraginaceae s.l., se caracteriza por presentar inclusiones celulares a manera de arena cristalina. En el presente estudio estas inclusiones se observaron en V. linnaei y en V. bullata, mientras que V. inermis no presentó sales cristalinas de ningún tipo. Gottwald (1983), señala que las inclusiones cristalinas poseen un gran valor taxonómico, tanto así que logró separar a todas las secciones de Boraginaceae s.l. (clasificación de Johnston 1930) utilizando este carácter. Sin embargo, para Taroda (1984), Gottwald enfatizó demasiado en las inclusiones cristalinas en lugar de estudiar las correlaciones involucrando diferentes caracteres y no tomarlos como parámetros aislados. La experiencia con las especies estudiadas nos dice, que estas inclusiones cristalinas pudieran tener un valor taxonómico, ya que se observó diferencia en la presencia, abundancia y distribución de las mismas dentro de la hoja.
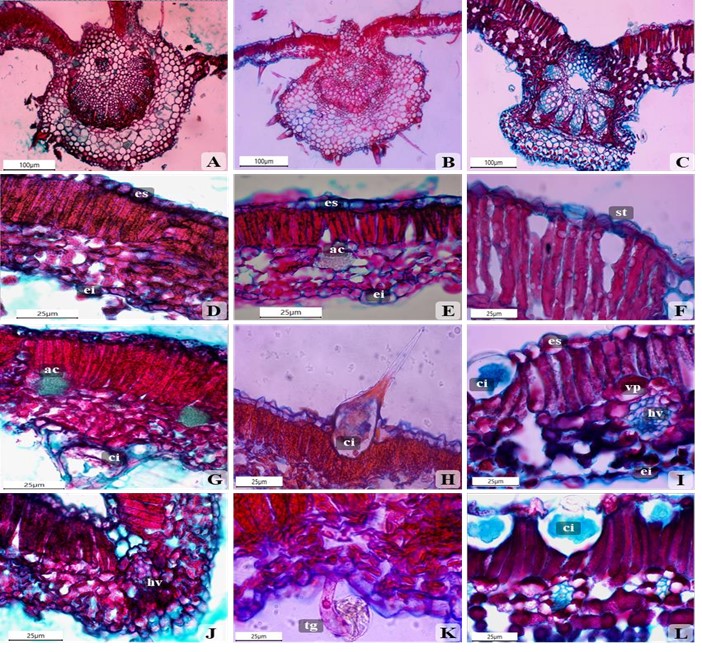
Fig. 2
Secciones transversales de la lámina foliar de Varronia bullata (A, D, G, J), Varronia linnaei (B, E, H, K) y Varronia inermis (C, F, I, L). es = Epidermis adaxial, ei = Epidermis abaxial, ac = Arena cristalina, st = Estoma, ci = Cistolito, hv = Haz vascular, vp = Células de la vaina de haz, tg = Tricoma glandular
Comparación de la superficie foliar entre las especies de Varronia.
En relación con la superficie foliar, los resultados muestran que se encontró similitud en el tipo de estomas (anomocítico), la presencia de tricomas simples en ambas superficies, y la presencia de tricomas simples y glandulares en la epidermis abaxial, mientras que caracteres como la distribución de los estomas en la hoja y los tipos de tricomas glandulares, difieren entre las especies e incluso entre las poblaciones de una misma especie.
Epidermis (en vista superficial)
La epidermis adaxial presenta células poligonales con paredes que van desde rectas a onduladas. También se observaron células epidérmicas de contorno poligonal dispuestas radialmente a modo de roseta alrededor de los tricomas simples. El número y tamaño de estas células epidérmicas varía para cada especie (Fig. 3A, 3B, 3C, 3D, 3H, 3I).
Tipo de tricomas
En todas las especies se observaron tricomas simples y glandulares. Los tricomas simples poseen paredes papilosas (Fig. 3G), se presentan en ambas superficies y pueden ser de dos tipos: localizados directamente en la superficie de la epidermis (Fig. 4E, 4F, 4H) o localizados sobre elevaciones de células epidérmicas (Fig. 3A, 4A, 4G). En la mayoría de los casos, los tricomas simples de la epidermis adaxial presentaron bases cistolíticas (Fig. 3B, 3C, 3D).
Los tricomas glandulares se localizan en la epidermis abaxial, son uniseriados, tienen una base unicelular; un pie corto unicelular. Estos tricomas pueden ser de dos tipos: con la cabeza redonda (Fig. 5D, 5E, 5F, 5G) o con la cabeza ovalada (Fig. 3E, 5H). Estos últimos, solo fueron visibles en V. inermis. Los tricomas simples con paredes papilosas observados en este estudio, son muy parecidos a los tricomas con paredes papilosas sobre bases glabras, encontrados por Mehrabian et, al. (2014) en las secciones Protonosma, Podonosma y Onosma (subsección Haplotricha) del género Onosma de la familia Boraginaceae. La aparición de tricomas simples con o sin bases cistolíticas y tricomas glandulares concuerda con la descripción realizada por Estrada (1995) para el género Varronia. A pesar de esto, nuestros resultados no concuerdan la descripción de los tricomas simples de este autor para V. linnaei, donde señala que la superficie adaxial en esta especie presenta tricomas simples junto con tricomas cistolíticos reducidos, por nuestra parte, los tricomas de esta superficie eran simples con bases cistolíticas, relativamente cortos, pero nunca reducidos.
En el mismo orden de ideas, Estrada (1995) también describe el indumento de V. bullata, y manifiesta que en la superficie adaxial se presentan tricomas simples con bases cistolíticas y en la superficie abaxial, tricomas simples adpresos junto con tricomas glandulares, tal como muestran nuestros resultados.
En V. inermis se encontraron tricomas cistolíticos reducidos en la superficie adaxial, como los que describió Estrada (1995) para V. linnaei y muy parecidos a los que encontraron Ventrella y Marinho (2008) en Cordia verbenacea. Para el caso de V. inermis, en el presente estudio se describe por primera vez el indumento de esta especie, siendo un aporte al conocimiento del género Varronia.
Densidad de los tricomas
Tomando en cuenta la clasificación del indumento de acuerdo a la densidad de tricomas propuesta por Barykina y Alyonkin (2016), para algunas de las especies de Boraginaceae, se manifiesta que las tres especies poseen un indumento muy denso en la epidermis abaxial, mientras que en la epidermis adaxial V. bullata presenta un indumento denso y el resto de las especies un indumento muy denso.
Detalles de las diferencias en el indumento para cada especie de Varronia se muestran en las Figuras 3E, 3F, 4A, 4B, 4C Y 4D.
Posición y tipo de estomas
En todas las especies estudiadas, los estomas son del tipo anomocítico (Fig. 3H, 3I,4I). En cuanto a su posición, se ubican en la epidermis abaxial caracterizando a la hoja como hipostomática, exceptuando a las poblaciones de Alanje y Limones de V. inermis, donde los estomas se ubican en ambas epidermis, lo que hace que la hoja sea definida como anfiestomática (Fig. 3B, 3C, 4I).
Las especies de Varronia estudiadas presentaron estomas anomocíticos, tal como mencionan Watson (1992) y Dasti (2003) en sus estudios con este género. Tomando en cuenta la distribución de los estomas, Metcalfe y Chalk (1950) definen la lámina foliar de Cordia (incluido Varronia) como anfiestomática. Años más tarde, Watson (1992), manifiesta que en el género Varronia las hojas pueden ser tanto hipostomáticas como anfiestomáticas. Nuestros resultados apoyan a Watson (1992), ya que observamos hojas hipostomáticas en V. linnaei, V. bullata y V. inermis (población de Río Guanábano) y hojas anfiestomáticas en las poblaciones de V. inermis de Limones y Alanje. Además, Tölke et al. (2013), reportaron hojas hipostomáticas en V. leucocephala y hojas anfiestomáticas en V. globosa.
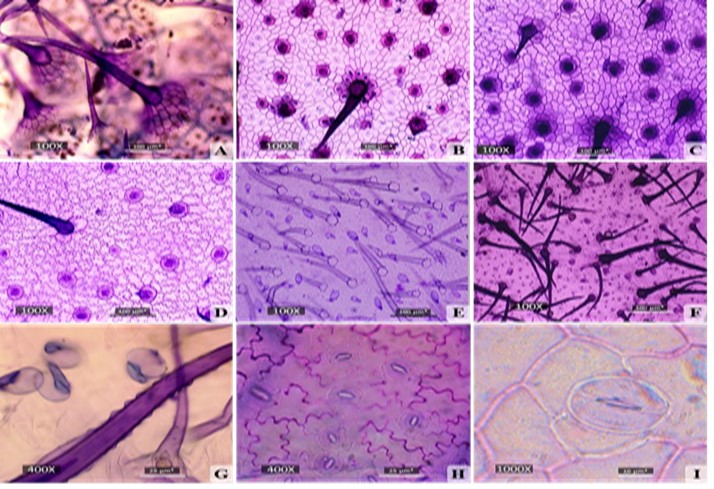
Fig. 3
Vista superficial de la lámina foliar de las especies de Varronia. A) Epidermis adaxial de V. bullata. B) Epidermis adaxial de V. inermis (población de Alanje). C) Epidermis adaxial de V. inermis (población de Alanje). D) Epidermis adaxial de V. inermis (población de Río Guanábano). E) Tricomas simples y glandulares en la epidermis abaxial de V. inermis. F) Tricomas simples y glandulares en la epidermis abaxial de V. linnaei. G) Papilas en los tricomas simples de V. bullata. H) Estomas en la epidermis abaxial de V. inermis. I) Estomas en la epidermis abaxial de V. linnaei
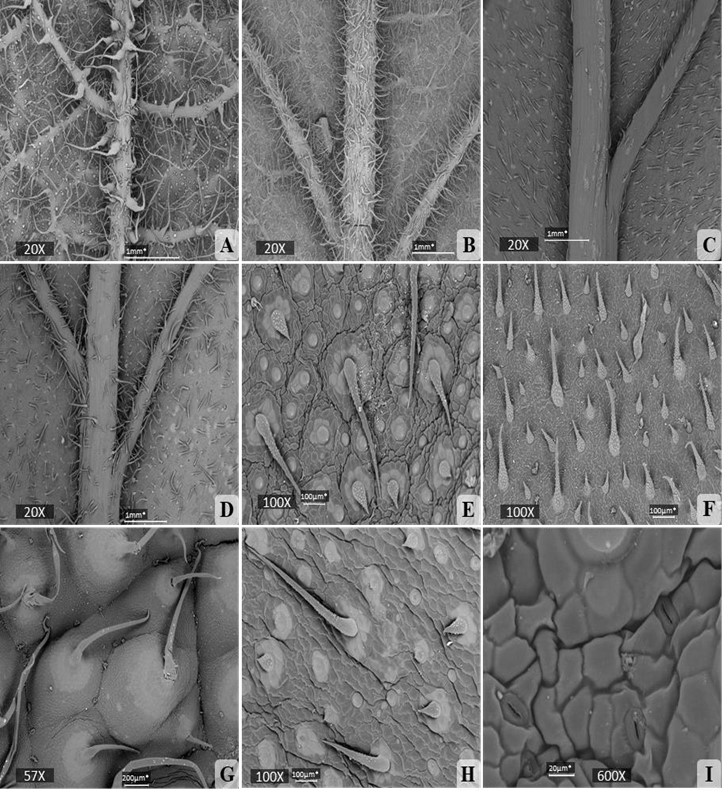
Fig. 4
Microfotografías tomadas con el MEB de la lámina foliar de las especies de Varronia. A) Epidermis abaxial de V. bullata. B) Epidermis abaxial de V. linnaei. C) Epidermis abaxial de V. inermis (población Río Guanábano). D) Epidermis abaxial de V. inermis (población Limones). E) Epidermis adaxial de V. inermis (población Río Guanábano). F) Epidermis adaxial de V. linnaei. G) Epidermis adaxial de V. bullata. H) Epidermis adaxial de V. inermis (población Limones). I) Estomas en la epidermis adaxial de V. inermis (población Limones)
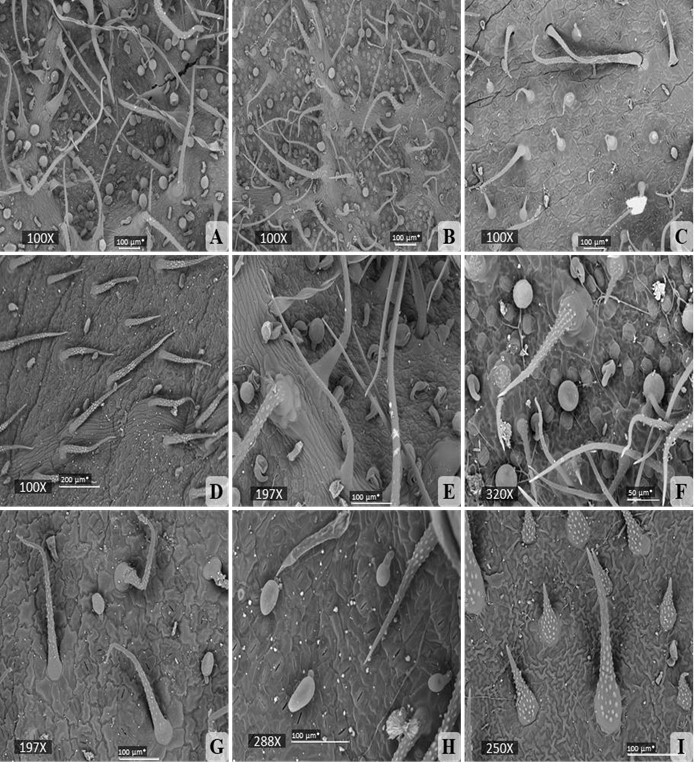
Fig. 5
Microfotografías tomadas con el MEB representando la densidad los tricomas de las especies de Varronia. A) Tricomas simples y glandulares en V. bullata. B) Tricomas simples y glandulares en V. linnaei. C) Tricomas simples y glandulares en V. inermis (población Limones). D) Tricomas simples con papilas y tricomas glandulares en V. inermis (población Río Guanábano). E) Tricomas simples con base pluricelular en forma de roseta junto a tricomas glandulares en V. bullata. F) Tricomas simples con base pluricelular en forma de roseta junto a tricomas glandulares en V. linnaei. G) Tricomas simples con paredes papilosas y tricomas glandulares en V. inermis. H) Tricomas glandulares redondeados y ovalados en V. inermis. I) Tricomas simples con paredes papilosas en V. linnaei
Análisis de datos
A partir de la matriz de datos cuantitativos, se realizó un análisis discriminante, en el cual se utilizaron 226 casos y se introdujeron 11 variables predictoras para desarrollar un modelo que discriminara entre los 3 niveles de especie.
Para separar los datos de una manera óptima mediante la combinación lineal de las variables, se obtuvieron las dos funciones discriminantes (representadas por el eje 1 y el eje 2). La primera función discriminante poseía un porcentaje de varianza relativo de 75,79 %, mientras que la segunda función discriminante sólo era capaz de explicar 24,08 % de la varianza de los datos. Sin embargo, el valor-p de ambas funciones discriminantes fue inferior a 0,05; con lo cual ambas funciones discriminantes eran estadísticamente significativas con un 95% de confianza. Por último, la correlación canónica de la primera función discriminante estaba más próxima a 1, lo que indicaba que esta función fue capaz de separar mucho mejor los datos que la segunda.
Para la primera función discriminante, la variable “longitud de tricomas simples en la epidermis adaxial” posee el coeficiente de mayor magnitud (0,93), mientras que para la segunda función discriminante el valor más alto está representado por la “densidad de tricomas simples en la epidermis abaxial” con 0,72.
A partir de las dos funciones discriminantes resultantes, se graficaron las observaciones para cada una de las especies (Fig. 6). La imagen nos muestra cómo se separan las tres especies en estudio. Siendo V. bullata, la especie que queda más alejada de las otras especies. De esta forma se puede manifestar que es posible una separación de las tres especies del complejo Varronia bullata, utilizando las variables escogidas para el estudio.
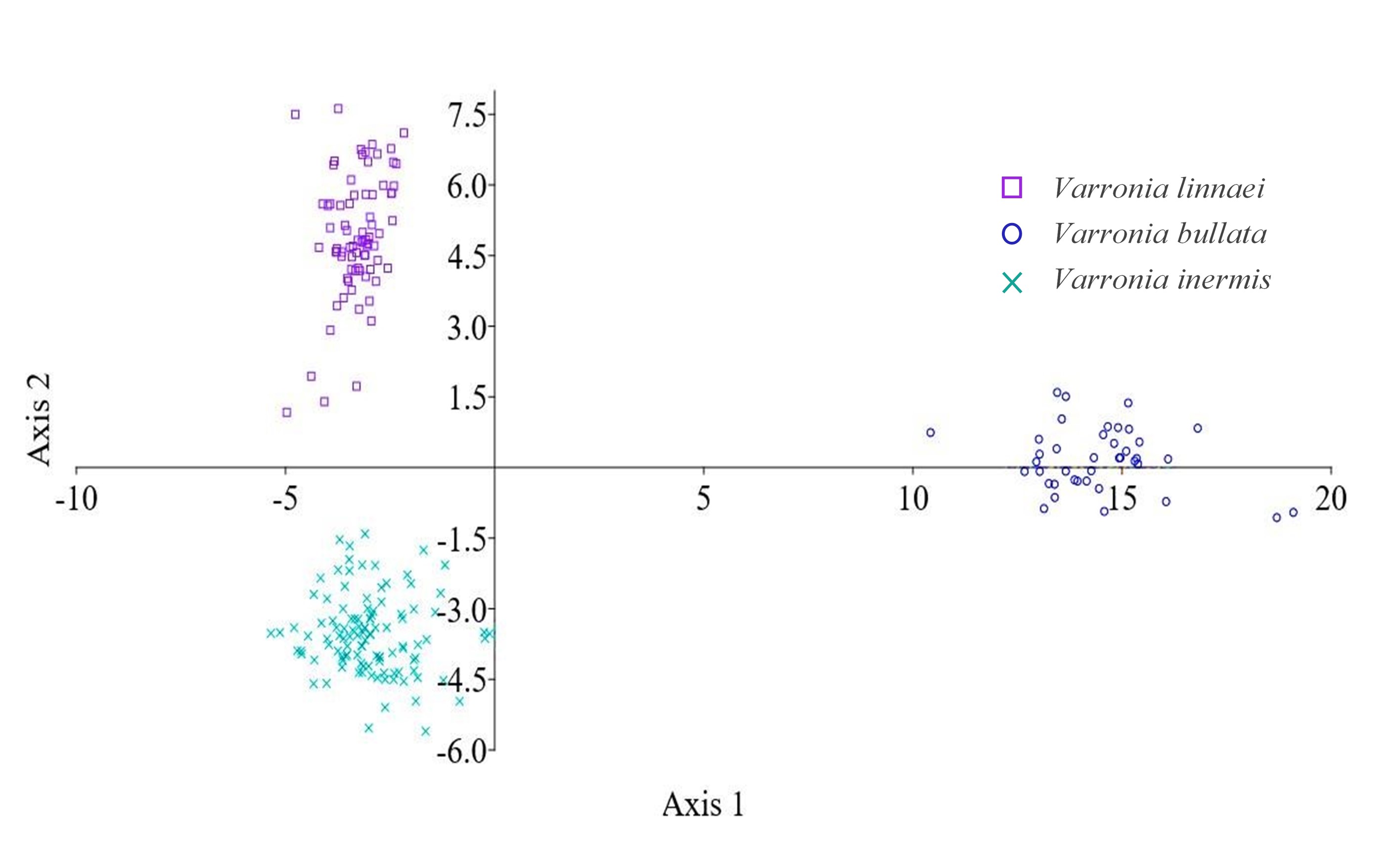
Fig. 6
Gráfico de dispersión. Representación de la separación de las especies de Varronia. utilizando las funciones discriminantes
Los resultados obtenidos en este estudio coinciden con los trabajos de Metcalfe y Chalk (1950), Watson (1992), Estrada (1995), Dasti (2003) y Tölke et al. (2013), quienes reportaron una epidermis uniestratificada, mesófilo dorsiventral, tricomas sobre bases cistolíticas y la presencia de inclusiones celulares para el género Varronia.
Vilela (1996), manifiesta que la anatomía en las plantas es considerada, junto con las características morfológicas, ecológicas y genéticas, como una valiosa herramienta para la clasificación de los distintos taxa. Si bien es cierto, las especies en estudio pueden ser definidas anatómicamente, hubo grandes rangos de variaciones intraespecíficas en parte de los caracteres estudiados, como lo fueron el grosor de la epidermis, el grosor del parénquima y la densidad estomática. Esto puede explicarse desde el punto de vista fisiológico, como sugiere Molina (2008), las plantas modifican su morfología a lo largo de las gradientes altitudinales como estrategia para mitigar las condiciones climáticas limitantes. Dentro de estas modificaciones, las más comunes son: el área foliar, grosor de las hojas y densidad de estomas. También hay variación en características y tamaño de las células. Debido a la altitud aumenta la radiación fotosintéticamente activa y UV, la presencia de filtros mecánicos, como pelos foliares o filtros fotoquímicos, como pigmentos protectores se ven aumentados. Por lo que, en las primeras etapas del desarrollo de las hojas, los tricomas provistos de polifenoles pueden tener una función protectora frente a los daños que pudieran causar las radiaciones UVB (Brownlee, 2001; Esaú, 1977). La pubescencia foliar podría actuar como primera barrera frente a la alta radiación solar (Molina, 2008). En términos generales, y sin haber tomado en cuenta ningún factor ambiental, nuestros resultados concuerdan con lo expuesto por estos autores, tal es el caso de las hojas de V. bullata las cuales se colectaron a la mayor altitud para este estudio (800 msnm), y fueron las más gruesas (epidermis superior + parénquima en empalizada + parénquima esponjoso + epidermis inferior), además de obtener los valores más altos para la densidad estomática.
Aunque los caracteres relacionados a variaciones ambientales representan parte importante en la definición de un taxón y nos permiten ver el grado de variación intraespecífica y la plasticidad fenotípica de cada especie; se concluye entonces, que la utilización con fines taxonómicos de caracteres variables a causa de las condiciones ambientales, podría llevar a una confusión mayor.
Aun cuando la estructura foliar en Varronia es similar para muchos caracteres, diferencias marcadas en la longitud de los tricomas simples, el tipo de tricomas, la densidad estomática y la densidad de los tricomas simples y glandulares en la epidermis abaxial, así como la presencia o ausencia de arena cristalina, nos permitieron distinguir entre las especies en estudio.
En otro contexto, anatómicamente, el ejemplar que es diferente al resto de su especie, especialmente en el tipo de hoja en relación a la distribución estomática es el proveniente de la población de Río Guanábano (V. inermis). Evidentemente, el principal factor que rige las homologías moleculares es genético, mientras que, para las variables anatómicas y morfológicas, las restricciones ambientales juegan un papel muy importante (Vilela, 1996). Para este caso en particular, sería difícil asignar un rango taxonómico basándonos exclusivamente en caracteres morfoanatómicos, hace falta más estudios que permitan discernir con mayor precisión lo que sucede en esta población, con relación a las demás poblaciones de su misma especie.
Mediante este estudio se comprueba el potencial que posee la morfoanatomía foliar en la delimitación taxonómica del complejo Varronia bullata: V. inermis, V. linnaei y V. bullata- Esta metodología permitió separar las especies de este complejo utilizando características morfoanatómicas tales como la densidad de tricomas simples en la epidermis abaxial, la longitud de los tricomas simples en la epidermis adaxial, la densidad estomática y la densidad de tricomas glandulares en la epidermis abaxial.
Las especies del género Varronia estudiadas presentan similitudes en cuanto a los estomas anomocíticos, la epidermis uniestratificada, la aparición de tricomas simples sobre bases cistolíticas y tricomas glandulares unicelulares. En V. linnaei y V. bullata la lámina foliar es hipostomática, mientras que en V. inermis es hipostomática o anfiestomática, de igual forma, las dos primeras especies muestran sales cristalinas incluidas en el mesófilo y en V. inermis no se encontró ninguna evidencia para este carácter.
Las características relacionadas a los tricomas, como su forma, tamaño, distribución y densidad; así como la presencia de inclusiones cristalinas también juegan un papel importante en la separación taxonómica de las tres especies del complejo Varronia bullata.
A la Vicerrectoría de Asuntos Estudiantiles de la Universidad de Panamá por financiar parte de la investigación y al Herbario de la Universidad de Panamá por el uso de sus instalaciones para el procesamiento de las muestras.
Fig. 1
Localidades de colecta de las especies estudiadas
Fig. 2
Secciones transversales de la lámina foliar de Varronia bullata (A, D, G, J), Varronia linnaei (B, E, H, K) y Varronia inermis (C, F, I, L). es = Epidermis adaxial, ei = Epidermis abaxial, ac = Arena cristalina, st = Estoma, ci = Cistolito, hv = Haz vascular, vp = Células de la vaina de haz, tg = Tricoma glandular
Fig. 3
Vista superficial de la lámina foliar de las especies de Varronia. A) Epidermis adaxial de V. bullata. B) Epidermis adaxial de V. inermis (población de Alanje). C) Epidermis adaxial de V. inermis (población de Alanje). D) Epidermis adaxial de V. inermis (población de Río Guanábano). E) Tricomas simples y glandulares en la epidermis abaxial de V. inermis. F) Tricomas simples y glandulares en la epidermis abaxial de V. linnaei. G) Papilas en los tricomas simples de V. bullata. H) Estomas en la epidermis abaxial de V. inermis. I) Estomas en la epidermis abaxial de V. linnaei
Fig. 4
Microfotografías tomadas con el MEB de la lámina foliar de las especies de Varronia. A) Epidermis abaxial de V. bullata. B) Epidermis abaxial de V. linnaei. C) Epidermis abaxial de V. inermis (población Río Guanábano). D) Epidermis abaxial de V. inermis (población Limones). E) Epidermis adaxial de V. inermis (población Río Guanábano). F) Epidermis adaxial de V. linnaei. G) Epidermis adaxial de V. bullata. H) Epidermis adaxial de V. inermis (población Limones). I) Estomas en la epidermis adaxial de V. inermis (población Limones)
Fig. 5
Microfotografías tomadas con el MEB representando la densidad los tricomas de las especies de Varronia. A) Tricomas simples y glandulares en V. bullata. B) Tricomas simples y glandulares en V. linnaei. C) Tricomas simples y glandulares en V. inermis (población Limones). D) Tricomas simples con papilas y tricomas glandulares en V. inermis (población Río Guanábano). E) Tricomas simples con base pluricelular en forma de roseta junto a tricomas glandulares en V. bullata. F) Tricomas simples con base pluricelular en forma de roseta junto a tricomas glandulares en V. linnaei. G) Tricomas simples con paredes papilosas y tricomas glandulares en V. inermis. H) Tricomas glandulares redondeados y ovalados en V. inermis. I) Tricomas simples con paredes papilosas en V. linnaei
Fig. 6
Gráfico de dispersión. Representación de la separación de las especies de Varronia. utilizando las funciones discriminantes


