2020
31
12102020
23112020
 José Isaac Norato jose.norato@up.ac.pa
José Isaac Norato jose.norato@up.ac.pa
Universidad de Panamá,, Panamá
Edil Enrique Araúz edil.arauz@up.ac.pa
Universidad de Panamá, , Panamá
Resumen: La bioestimulación hematopoyética (BH) mineral (Fe, Cu y Co) y vitamínica (B1, B12) fue estudiada en las primeras 16 semanas después del parto, para evaluar su posible impacto sobre el perfil hematimétrico y celular sanguíneo en vacas Holstein en producción en una finca lechera grado A, ubicada a 1160 metros de altitud en el clima tropical húmedo. Se utilizó cuatro vacas Holstein de una, dos, tres y cuatro lactaciones; asignando al azar dos vacas de cada lactación para aplicar la solución estéril hemopoyética [gluconato de hierro (10 miligramos (mg) / mililitro (mL, gluconato de cobre (10 mg/mL, gluconato de cobalto (10 mg/mL, tiamina (100 mg/mL y cianocobalamina (20 mg/mL) y dos vacas para solución salina fisiológica; en las cuales, se administró 15 mL IM a los 10; 40 y 70 días postpartales. La BH aumentó el conteo de eritrocitos y la concentración corpuscular de hemoglobina (P<0,0001) en la 1ra lactación en +7,41 y +8,97% y en la segunda lactación en + 6,17% en los eritrocitos y +6.21% en la hemoglobina; pero en la 3ra y 4ta lactación no hubo diferencias estadísticas (P>0,05); aunque superiores al control. La BH aumentó las plaquetas (P<0,0528), los glóbulos blancos (P<0,0147) y los neutrófilos (P<0,0769); pero no modificó el hematocrito (P<0,05), los linfocitos (P>0,05) y los monocitos (P>0,05). La estimulación hemopoyética postpartal aumentó el perfil celular sanguíneo; indicando que el tejido mieloide fue activado en la citogénesis, maduración y liberación de eritrocitos, leucocitos, trombocitos y hemoglobina en la fase lactacional temprana en vacas Holstein bajo manejo lechero intensivo tropical.
Palabras clave: Hematopoyesis, hemoglobina, eritrocitos, leucocitos, trombocitos, linfocitos, Holstein, lactación.
Abstract: The hematopoietic biostimulation based on minerals (Fe, Cu and Co) and vitamins (B1 and B12) was studied across the first 16 weeks after parturition to evaluate its effects on blood indicators and cellular profiles in lactating Holstein cows in a grade A dairy farm at altitude of 1160 meters. Four lactating Holstein cows at one, two, three and four lactations were used. Two of them by lactation were randomly assigned to a treatment of sterile solution containing [Iron gluconate (10 mg/mL), copper gluconate (10 mg/mL), cobalt gluconate (10 mg/mL), thiamin (100 mg/mL) and cyanocobalamin (20 mg/mL)] and the other two cows by lactation received physiological saline solution (control). Animals were injected for treatment and control 15 ml IM at 10, 40 and 70 days after parturition. The hematopoietic stimulation significantly increased red blood cell count and mean corpuscular hemoglobin concentration (P<.0001) at the 1st and 2nd lactation; showing differences of +7,41% and +8,97%; while, mean corpuscular hemoglobin concentration showed differences +6,17% and +6,21%. However, animals in the 3rd and 4th lactation did not show statistically differences (P>0,05), but were higher. The stimulation increased platelets (P<0,05), White blood cells (P<.01) and neutrophils (P<0,10); however, the accumulated red cell volume, lymphocyte and monocyte were not modified (P>0,05). The hematopoietic stimulation increased some blood cells; indicating that myeloid tissue was activated on its cytogenesis, maturation and liberation of erythrocytes, leukocytes and platelets; as well as hemoglobin across the first third of lactation in Holstein cows under intensive dairy management in the tropics.
Keywords: Haematopoyesis, hemoglobin, erythrocyte, leukocyte, platelets, lymphocyte, Holstein, lactation.
INFLUENCIA DE LA BIOESTIMULACIÓN HEMATOPOYÉTICA MINERAL (Fe, Cu y Co) Y VITAMÍNICA (B1, B12) SOBRE EL PERFIL SANGUÍNEO POSTPARTAL EN VACAS HOLSTEIN EN LACTACIÓN
INFLUENCE OF MINERAL (Fe, Cu AND Co) AND VITAMIN (B1 AND B12) HAEMATOPOIETIC BIOSTIMULATION ON POSTPARTUM BLOOD CELLULAR PROFILES IN LACTATING HOLSTEIN COWS
José Isaac Norato jose.norato@up.ac.pa
Edil Enrique Araúz edil.arauz@up.ac.pa
Recepción: 12 Octubre 2020
Aprobación: 23 Noviembre 2020

Los primeros noventa días (d) después del parto son de vital importancia para las vacas lecheras, ya que este periodo exige gran cantidad de nutrientes; principalmente energía para cubrir las necesidades de mantenimiento y las funciones de producción. El parto, es el punto que marca el inicio de la lactación y es donde ocurre la mayor pérdida de sangre a causa del desprendimiento de las membranas placentarias, sin mencionar, las pérdidas diarias causadas por los hemoparásitos que afectan el perfil sanguíneo; ocasionado una mayor respuesta y presión sobre el tejido mieloide y los demás órganos hematopoyéticos (Dukes y Swenson, 1993; Fraser et al., 1993; Luna y Roldan, 2013; Morrow, 1983).
Luna y Roldan (2013) demostraron que los valores del hierro (Fe) y cobre (Cu) en sangre tienden a disminuir en el parto y el inicio de la fase de lactación; hecho que es crítico ya que ambos son minerales demandados por la médula ósea para la formación de hb. Esta proteína contiene en su interior cuatro átomos de Fe (Hurley y Doane, 1989). El 95% del Cu del organismo está en forma de ceruloplasmina; proteína que facilitar la absorción y el transporte del Fe al tejido hematopoyético (Underwood y Suttle, 2003); específicamente al tejido mieloide responsable de la eritrogénesis y de la hemoglobinopoiesis (Cunningham y Klein, 2009).
El cobalto (Co) se describe junto a la vitamina B12 o cianocobalamina debido a su relación con el metabolismo energético y de los lípidos (Kraft y Dürr, 2005). El Co es la materia prima utilizada por los microorganismos ruminales para sintetizar la cobalamina. Cerca de 4,4% del Co está contenido en la molécula de cianocobalamina, sin embargo, el rumiante hace uso extremadamente ineficiente de esta vitamina hidrosoluble (Underwood, 1981). La tiamina o vitamina B1, también es hidrosoluble y está fuertemente ligada al metabolismo energético (Butler y Smith, 1989), al igual que la cobalamina, pero ambas actúan de forma distinta y son necesarias en la obtención de energía química.
La tiamina, modula el paso del ácido pirúvico resultante del glicólisis hacia la acetil-Co A, la cual, entra al ciclo de Krebs para producir energía (Corbellini, 2000) en forma de Adenosin
Trifosfato (ATP). Sin embargo, la Adenosin-cobalamina (forma activa de la cianocobalamina), modula el paso del Succinil-Co A a propionato; además interviene en la formación de glucosa hepática o gluconeogenesis (Underwood y Suttle, 2003).
La respuesta hematopoyética influye primeramente en el metabolismo de carbohidratos, lípidos y proteínas, la cual, requieren del aporte de nutrientes para el tejido mieloide y linfoide (Swanson, 1989); tales como: el Fe, Cu, Co, B1 y B12. Estos micro factores nutricionales influyen en los demás sectores hematopoyéticos (Hígado, Bazo). Los minerales y vitaminas tipo B son necesarios para producir y reponer las células sanguíneas no funcionales que son eliminadas en el bazo y el hígado. La vida media de un eritrocito es de 120 d, plaquetas hasta 10 d, mientras que los leucocitos varían desde unos pocos minutos hasta varios d (Weiss et al., 2010). Los efectos multidimensionales en tiempo de la BH parenteral, se ilustran en la Figura 1.
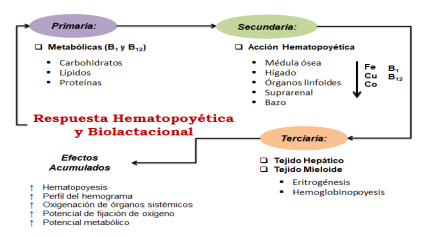
El tejido mieloide está en la medula ósea y ocupa la cavidad interior de los huesos largos y en las zonas trabeculares de los huesos planos. Este tejido experimenta cambios con la edad y su función no es igual en animales jóvenes en comparación con los adultos; ya que se estima que la receptividad de nutrientes hematopoyéticos difiere con la edad del animal (Dukes y Swenson,
1993) y se intensifica en animales recién nacidos a causa de la rápida proliferación de eritrocitos; los cuales, ejercen una fuerte demanda al tejido eritropoiético (Underwood y Suttle, 2003).
La hematopoyesis es la actividad citogénica y de maduración que realiza el tejido mieloide y los ganglios linfáticos en los rumiantes adultos (Dukes y Swenson, 1993); sin embargo, participan otros órganos como el estómago (producción de ácido clorhídrico y capacitación del hierro), el intestino (fijación y transporte del hierro), el bazo (destrucción de células circulantes no funcionales), los riñones (producción de eritropoyetina) y el hígado (almacenamiento de minerales, degradación de la hemoglobina y destrucción de células circulantes); los cuales forman parte del sistema hematopoyético complementario o de apoyo a la medula ósea y a los ganglios linfáticos para la producción de las células sanguíneas (Cunningham y Klein, 2009 ).
Los microfactores minerales participan en diversas formas en la hematopoyesis; discándose al Fe como componente del grupo hemo que forma parte de hb y de la mioglobina. Otras funciones enzimáticas del Fe como constituyente forman parte de la enzima de Transporte de electrones, de la oxidasa citocrómica, de la ferredoxina, de la enzima mieloperoxidasa, de la catalasa y del citocromo 450 (NRC, 2001).
El Cu es esencial para la absorción y el transporte del Fe; así como para la síntesis de hb y para la producción de la enzima dismutasa superoxide. Esta enzima protege las células de los efectos tóxicos de los derivados metabólicos del oxígeno y esencial para la fagocitosis (NRC, 2001). El cobalto es necesario para prevenir la anemia y para la formación bacteriana de la cianocobalamina en el rumen (NRC, 1989).
Las necesidades de minerales y vitamínicas han sido descritas por la NRC (1989,2001) para un bovino (Bos taurus) adulto; indicando que el Fe es requerido entre 60 a 80 mg/d; confirmado por otras fuentes (Kraft y Dürr, 2005). El requerimiento de Cu para vacas en lactación es de 10 partes por millón (ppm) (NRC, 2001); mientras que el Co solamente es necesario entre 0,04 a 0,06 mg/kilogramo (kg) de materia seca (Underwood, 1981; NRC, 1989).
Ubicación del estudio, área geográfica y entorno micro climático
El estudio fue desarrollado entre marzo y noviembre del 2018 en la cuenca lechera de Bugaba, específicamente en la comunidad de Bijao, vía Volcán, en la Finca lechera grado A los Milagros, a una elevación de 1160 metros sobre el nivel del mar (m.s.n.m.), temperatura en época lluviosa mínima de 18°C y máxima de 31,1°C en campo (Araúz et al., 2010).
El índice de temperatura humedad (ITH) mínimo fue calculado en 63,4 y el máximo en 83,4; el ITH promedio diario fue de 72,6 por lo tanto, el grado de tensión calórica microambiental a los que estuvieron sometidos los animales fue ligero a moderado durante tres horas dado la altitud de la finca experimental y en consecuencia dicho microambiente no representó un reto fisiológico extremo para las unidades experimentales (Araúz et al., 2011; McDowell, 1981).
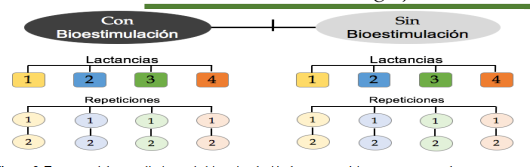
Se utilizó un total de 16 vacas Holstein, distribuidas en la 1era, 2da, 3era y 4ta lactación; formando cuatro grupos de cuatro animales; tal como se ilustra en la Figura 2. Todas las vacas fueron previamente evaluadas por su historial de salud, haciendo énfasis en sus cuatro cuartos funcionales, ausencia de mastitis, ausencia de antibióticos y que no hubiesen presentado problemas al parto ni enfermedades metabólicas (Fiebre de leche, acetonemia, síndrome de vaca gorda y síndrome de la vaca delgada). Al grupo bioestimulado, se le aplicó, a cada vaca, por vía intramuscular (IM) 15 mL de una solución comercial estéril a base de gluconato de Fe (10mg/mL), gluconato de Cu (10mg/mL), gluconato de Co (10mg/mL), tiamina (100mg/mL) y cianocobalamina (20 mg/mL); en los D 10; 40 y 70 después del parto.
El producto utilizado, fue una solución llamada anti anémica, cuya procedencia es Argentina y cuenta con el número de registro sanitario en Panamá RF-6809-14, vigente hasta el 20 de marzo de 2024 (27). Cada aplicación de este producto aportó al animal 150 mg de Fe, 150 mg de Cu, 150mg de Co, 1500 mg de B1 y 300 mg de B12. Al grupo bioestimulado se le aplicó 15 mL de la solución Antianémica y el grupo control se le administró Solución Salina Fisiológica igualmente en los d 10; 40 y 70 después del parto.
Las 16 vacas en total fueron muestreadas cada 15 d; extrayendo una muestra de sangre de 3,5 mL de la vena coccígea y luego colocada en tubos de ensayo con anticoagulante, ácido etilen diamino tetracético (EDTA). Todas las muestras colectadas fueron llevadas al laboratorio de Cooleche R.L, ubicado en el distrito de Bugaba, Provincia de Chiriquí, Panamá. El análisis hematimétrico fue realizado por hematología computarizada.
Los parámetros para caracterizar el perfil hemático fueron el ht o volumen celular eritrocítico aglomerado (vcea) (%), hb (gramos (g)/decilitro (dL)), concentración corpuscular media (ccm) de la hb (g/dL), eritrocitos (millones/mL), leucocitos (miles/mL), trombocitos (miles/mL), linfocitos (miles/mL) y monocitos (miles/mL). Los análisis de la sangre fueron realizados en el
laboratorio Especializado de la Cooperativa de Productores de Leche (Cooleche R. L), ubicado en la Concepción, Bugaba, Provincia de Chiriquí, Panamá.
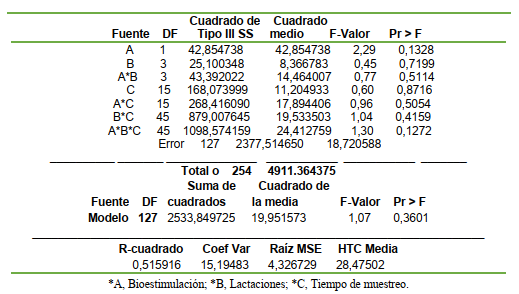
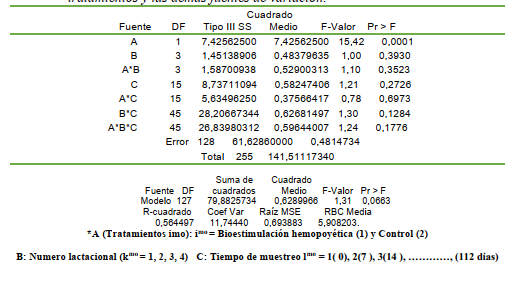
El experimento fue estructurado y analizado en base al Diseño Arreglo Factorial, incluyendo los tratamientos (Ai = 1(Bioestimulación) y 2 (control), el número lactacional kth [1ra, 2da, 3ra y 4ta lactación] y Cl el muestreo sanguíneo semanal (lmo = 0; 7; 14; 21; 28; 35; 42; 49; 56; 63; 70; 77; 84; 91; 98; y 105 d). El modelo lineal aditivo fue el correspondiente al arreglo factorial para tres factores de variación; descrito como:
Yijkl = µ + Ai + B(k) + (AB)ik + Ci + (AC)il + (BC)kl + (ABC)ikl + ℮(ijk) donde:
Yijkl es el conteo de células sanguíneas (eritrocitos, leucocitos, plaquetas, linfocitos, monocitos), hb y el ht. Los datos fueron analizados con el programa SAS (2002) según los procedimientos indicados por Herrera y Barreras (2000) y los procedimientos de las medias, la significancia y la tendencia genérica se realizaron según Gill (1978; SAS, 2002)
Volumen celular eritrocítico aglomerado o hematocrito (%)
El ht representa el volumen celular proporcional de los eritrocitos acumulados por centrifugación generado por unas 40,000 revoluciones aplicadas en cinco minutos (min) (Duran et al., 2005). El referente hematológico del ht en bovinos oscila entre 21 y 30% (33), 28 a 38 (33) y de 22 a 33% (Fraser et al., 1993). Los límites del ht para los bovinos es de 26 a 46% según Duncan y Prasse (1986) y la media es de 35% (PFIZER, 1998); indicando la proporción de los eritrocitos en el volumen de sangre por cada 100 mL.
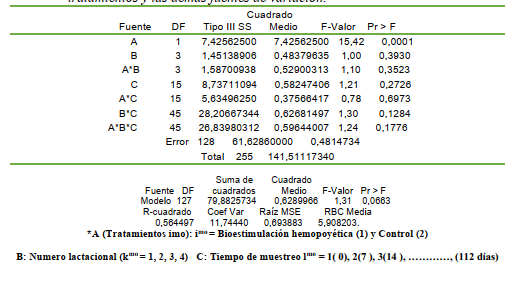
El análisis de varianza mostró que el ht no fue diferente entre los tratamientos (P>0,05), lactaciones (P>0,05) y entre las semanas (P>0,05) y no se observó la interacción entre los factores principales de variación (TABLA 1).
Las medias del vcea fueron generadas para las cuatro lactaciones evaluadas distinguiendo el tratamiento y el control para establecer los diferenciales proporcionales y relativos dentro de cada lactación como método alternativo. En términos relativos, se observó que las medias del ht en los animales bioestimuladas (Fe, Cu y Co) con los minerales y las vitaminas B1 y B2 fueron ligeramente superiores ante el control.
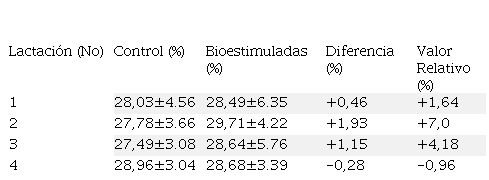
El ht venoso del grupo bioestimulado y control fue normal según Wood y Quiroz-Rocha (2010), Fraser et al. (1993) y George et al. (2010). En la segunda lactación se obtuvo un incremento de
7% al comparar la bioestimulación con el grupo control; a pesar de que estas medias no fueron diferentes estadísticamente (P>0,05). La inclusión del ht fue sustentado debido a la relación que hay entre el volumen de los eritrocitos con el volumen plasmático; sin embargo, la respuesta ante la BH fue traslada al conteo celular y a la concentración de hb; ya que las proporciones del volumen eritrocítico y plasmático están relacionadas a la respuesta hemopoyética.
Los glóbulos rojos y la hb son los responsables del transporte de los gases (O2, CO2) en la sangre y participan de la amortiguación del pH en el fluido sanguíneo como complemento fisiológico a partir de la oxihemoglobina y carboxihemoglobina con actividad ácida y alcalina (Dukes y Swenson, 1993; Wood y Quiroz-Rocha, 2010). La función primordial de los eritrocitos es transportar el O2 y el CO2 mediante la acción ligando de la hb; permitiendo la oxigenación celular y la detoxificación metabólica celular bajo los principios de las leyes de la difusión de los gases en los fluidos corporales (Duran et al., 2005; Guyton y Hall, 2012).
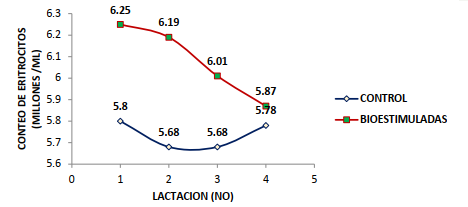
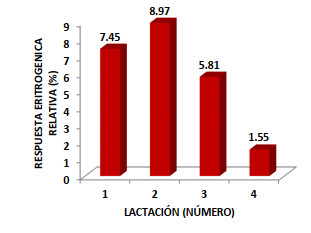
El conteo de los hematíes fue diferente entre el grupo de las vacas bioestimuladas y el grupo control (P<0,0001); ya que el rango normal se encuentra entre 5 y 7 millones/mL (PFIZER, 1998); aunque Fraser et al. (1993) indican que los límites normales son de 5 a 10 millones/mL. La BH mineral y vitamínica estuvo asociada con un mayor conteo de eritrocitos en los cuatros lactaciones evaluadas; evidenciando una respuesta positiva del tejido mieloide en el primer tercio lactacional (Figura 1). La respuesta eritrogénica se redujo al aumentar el número lactacional; destacando la influencia colateral de la edad y el historial lactacional en el proceso de la eritrogenesis vía la doble estimulación nutricional parenteral postpartal temprana a base de los minerales Fe, Cu y Co más tiamina y rivoflavina.
La BH estuvo asociada con un incremento en el conteo de los glóbulos rojos; que fue superior en las vacas de primera y segunda lactación, las cuales mostraron una elevación eritrocitica de 1,30X y 1,55X superiores al diferencial de los glóbulos rojos para la tercera lactación; mientras la respuesta eritropoyética en comparación con las vacas de cuarta lactación fue 4,77X (1ra lactación), 5,67X (2da lactación) y 3,17X (3ra lactación) en el periodo de 112 d luego de la exposición al proceso de la bioestimulación mineral – vitamínica.
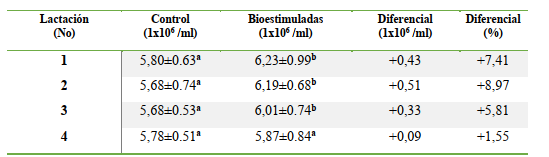
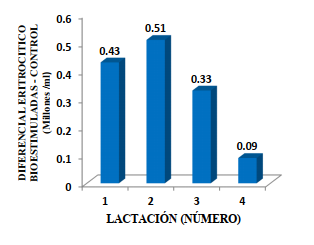
La acción simultánea de los minerales Fe, Cu y Co en conjunto con la tiamina y la cianocobalamina estuvo asociada con una respuesta eritrogénica positiva según el conteo de los glóbulos rojos y el diferencial entre la bioestimulación y el control; generando un valor relativo en las 1ra, 2da, 3ra y 4ta lactación; correspondiendo a +0.43, +0.51, +0.33 y +0.09 millones de eritrocitos/mL que representó un incremento en los glóbulos rojos circulantes de +7.45, +8.97,
+5.81 y +1.55% (Figuras 3, 4 y 5).
La disponibilidad de los microfactores minerales hematopoyéticos (Fe, Cu, Co) y de las vitaminas (B1, B12) facilitadoras de actividad celular en el tejido mieloide constituyen un entorno que facilita los procesos celulares de la diferenciación y maduración de las células mieloides incluyendo el eritroblasto, leucoblasto, megacarioblasto y monocitoblasto (Cunningham y Klein, 2009; Fraser et al., 1993; Hurley, W. L.; Doane, 1989).
El principio de la reserva y disponibilidad de los microfactores minerales Fe, Cu y Co constituyen el marco nutreceútico mineral limitante de la eritropoyesis, leucopoyesis y Hemoglobinopoiesis. En consecuencia, la National Research Council (NRC) ha establecido las necesidades nutricionales diarias para las vacas en lactación en el modelo lactacional fresco según la materia seca por d.

Aproximadamente el 60 % del Fe corporal esta unido a la hb (Underwood y Suttle, 2003); proteína que está constituida por cuatro unidades hemo unidas a cuatro globinas (10) y cada molécula de hemo contiene un átomo central de hierro en su estructura. El contenido de hb fue diferente entre los tratamientos (P<0,0001), entre las lactaciones (P<0,0094) y entre las semanas
del estudio (P<0,0513); así como las interacciones lactaciones x semanas (P<0,0001) y tratamientos x lactación x semanas (P<0,0001).
Fraser et al. (1993) indican que la concentración de hb normal en vacas adultas es de 11 g/100mL; sin embargo, otros autores consideran que puede oscilar entre 8 y 15 g/100mL (Contreras, 2012; Duncan, 1986; Duncan y Prasse, 1986; Fraser et al., 1993). La bioestimulación mineral y vitamínica de la hematopoyesis estuvo asociada con un incremento en la concentración de hb en todas las lactaciones; correspondiéndole +6,17, +6,21, +4,41 y +2,67% en la 1ra, 2da, 3ra y 4ta lactación (TABLA 7) Ello constituye un hallazgo relevante para un periodo de 112 d asociados con el tratamiento estimulatorio hemopoyética; lo que sugiere que el tejido mieloide se fue favorecido en la diferenciación, maduración y liberación de los glóbulos rojos y con ello en la síntesis de hb al mismo tiempo; mejorando el perfil eritrocítico y hemoglobínico que facilita la vehiculización, el transporte e intercambio de gases.

La bioestimulación mineral (Fe, Cu y Co) con tiamina y cianocobalamina aplicados a los 10; 40 y 70 d después del parto condujeron a un incremento en la concentración de hb en las cuatro lactaciones de forma sostenida en todo el periodo de exposición a la suplementación (P<0,0001); cuya superioridad total fue +4.65% (Figura 6). La respuesta hemoglobinopoyética se muestra después de la primera estimulación hematopoyética, que fue potenciada por la segunda aplicación con una respuesta aumentada a partir de la tercera intervención a los 70 d (10ma semana) después del parto. No obstante, en el grupo control se observó una respuesta compensatoria; iniciando desde la 3ra semana postpartal con una sustentabilidad hasta la 10ma semana de lactación postpartal con una zona continuada de depresión hemoglobínica.
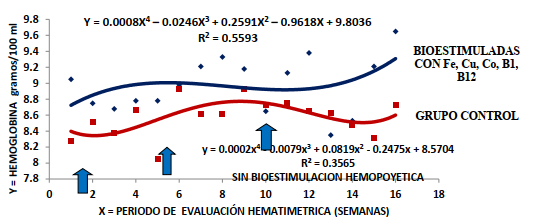
Finalmente, aunque en todas las lactaciones se produjo un aumento en la concentración de hb, la respuesta hemoglonipoietica se redujo desde la primera a la cuarta lactación; sugiriendo a su vez la importancia de contemplar la edad y el número lactacional para implementar la suplementación mineral y vitamínica hemopoyética en las vacas Holstein durante el primer tercio de la fase de producción lechera.
Los leucocitos son parte del protocolo diagnóstico sanguíneo para establecer la naturaleza de procesos infecciosos, parasitarios o traumatismos en los animales domésticos. La fisiología del parto conlleva pérdidas hemáticas sustanciales; incluyendo glóbulos rojos, leucocitos y trombocitos (Fraser et al., 1993;Nebel, 2011).
La reducción eritrocítica y leucocitaria del parto está acompañada previamente de la reducción de los anticuerpos circulantes antes del puerperio debido a la transferencia de inmunoglobulinas al tejido parenquimal mamario para la formación del calostro como parte de la calostrogénesis en la lactogénesis avanzada (Araúz y colaboradores, 2011). En consecuencia, ambas condiciones predisponen a la vaca durante la lactación temprana a sufrir infecciones, especialmente en el tracto reproductivo (Nebel, 2011) y por ende es esencial contemplar el perfil hematimétrico en el período postpartal para implementar las medidas nutraceúticas suplementarias para estimular la hematopoyesis. La estimulación hematopoyética con Fe, Cu y Co más tiamina y cianocobalamina no condujo a diferencias estadísticas con respecto al control (P>0,05) para los leucocitos; pero las vacas con bioestimulación mostraron medias del conteo leucocitario superiores al menos en las primeras tres lactaciones.
El conteo leucocitario mostró diferencias estadísticas al contemplar los tratamientos y el número lactacional (P<0,0147); indicando que la BH influyó sobre el conteo leucocitario en las primeras cuatro lactaciones; produciendo un aumento en el conteo de leucocitos en las primeras tres lactaciones; pero no en la cuarta lactación (TABLA 8). El conteo leucocitario normal en bovinos varía de 7 a 9,5 miles/mL (Pfizer, 1998); sin embargo, Fraser et al. (1993) señalan que el rango
normal es de 4 a 12 miles/mL En consecuencia, los animales control y los bioestimuladas mostraron un conteo leucocitario dentro de los límites normales. Sin embargo, puede observarse que los vacas bioestimuladas mostraron un conteo leucocitario circulante superior a los animales control (TABLA 9); evidenciando el incremento del conteo leucocitario y superando al grupo control en +29.56,
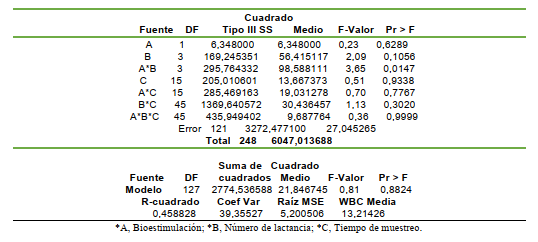

Las vacas bioestimuladas presentaron un conteo de leucocitos superior al grupo control, principalmente evidenciado en las dos primeras lactancias; indicando la influencia sobre el sistema mieloide, lo cual la leucopoyesis y por ende un mayor conteo leucocitario circulante; lo que representa una mayor capacidad para defender el organismo de los procesos infecciosos y traumáticos (Nebel, 2011). Es evidente que la BH influyó sobre el tejido mieloide, el cual, aportó
mayor cantidad de células formes al torrente sanguíneo, mejorando el perfil hemático de los animales bioestimuladas en comparación con el control.
La estimulación hemopoyética mineral y vitamínica postpartal aplicada en la etapa temprana de la lactación incrementó la hematopoyesis según los cambios en el conteo de eritrocitos, la concentración de la hb y el conteo de leucocitos en vacas Holstein sometidas al manejo lechero intensivo en condiciones tropicales.
El tejido mieloide fue potenciado con la aplicación del Fe, Cu y Co y tiamina y Cianocobalamina; estimulando los procesos celulares que acompañan la diferenciación, maduración y liberación de los eritrocitos y leucositos y por ende aumentando sus conteos a nivel circulante; con el acompañamiento del incremento en la concentración hb y el aumento de la capacidad de transporte de gases y la oxigenación sistémica derivada.
La aplicación múltiple de la estimulación mineral (Fe, Cu y Co) y vitamínica (B1, B12) temprano en la lactación potencia en forma aditiva la respuesta hematopoyética; aumentando perfil hematimétrico para los eritrocitos, hb hemoglobina y células leucocitarias totales; lo cual permite compensar la disminución celular circulante postpartal de estas células sanguíneas y beneficia en la protección celular leucocitaria y la oxigenación sistémica en la fase de mayor potencialidad lechera en las vacas Holstein.
Los autores declaran que el material presentado en este artículo científico no ha sido publicado en ninguna otra revista impresa o digital de carácter científico o técnico; que el contenido científico del artículo es el producto de la planificación y gestión de la investigación
debidamente coordinada por los autores y que la misma es originada por los autores dentro de la Universidad de Panamá propiamente. Los autores declaran que no existe diferencias en la conformación y/o contenido del documento científico como derivado de la investigación aplicada sobre la fisiología y hematología en la vaca lechera Holstein en lactación en las primeras cuatro lactaciones y en el primer tercio de la fase de la producción de leche. Los autores también declaran que este artículo ha sido generado para su consideración e inclusión en la Revista Científica de la Facultad de Ciencias Agropecuarias de la Universidad de Panamá.
Se les agradece muy especialmente a Empresa Lácteos Meyer al poner a nuestra disposición los animales y las facilidades físicas y el personal; así como por contribuir económicamente en los análisis de laboratorio y al nutricionista de la finca por su apoyo en esta investigación. A todos ustedes, nuestro eterno agradecimiento.


