

Dossier
Estudio ecomorfológico de la variación craneana en poblaciones prehistóricas de Perú
Ecomorphological study of the cranial variation in prehistoric populations from Peru
Estudo ecomorfológico da variação craniana em populações pré-históricas do Perú
Revista del Museo de La Plata
Universidad Nacional de La Plata, Argentina
ISSN: 2545-6377
Periodicidad: Semestral
vol. 5, núm. 1, 2020
Recepción: 01 Agosto 2019
Aprobación: 01 Diciembre 2019
Publicación: 01 Abril 2020

Resumen: El objetivo del presente trabajo es analizar la variación craneométrica en una muestra de individuos del Holoceno temprano/medio al tardío de Perú. Para esto, se registraron ocho medidas lineales en el esqueleto facial y bóveda craneana de 301 individuos procedentes de 19 sitios arqueológicos, los cuales se encuentran ubicados en cuatro biomas. Se exploró la variación en la forma y tamaño del cráneo entre individuos de distintos biomas y sitios arqueológicos mediante un Análisis de Componentes Principales y Análisis Discriminante. Complementariamente, se evaluó la afinidad morfológica entre las muestras mediante el cálculo de distancias de Mahalanobis y un análisis de clúster jerárquico de Ward. Asimismo, se estimó el efecto del bioma, sitio arqueológico, altitud, y cronología para explicar el patrón de variación morfológico observado a través de MANOVA. Los resultados mostraron que existen similitudes entre individuos de distintas cronologías; los individuos del Holoceno temprano/medio presentan gran variación morfológica; las muestras más recientes presentan diferencias en la longitud del cráneo, variables que describen la apertura nasal, pero también el subcomponente alveolar, lo cual a su vez se encuentra asociado a la diversidad de altitud y sitio arqueológico de procedencia. Se discuten los resultados en relación a trabajos previos que estudiaron muestras de Perú, y otras áreas de América del Sur. De esta manera se espera contribuir a la discusión de los debates actuales sobre el poblamiento de Sudamérica, especialmente en torno a las posibles adaptaciones locales.
Palabras clave: Evolución morfológica, Variación ecológica, Holoceno temprano a tardío, Andes Centrales.
Abstract: Ecomorphological study of the cranial variation in prehistoric populations from Peru. The aim of this paper is to analyze the craniometric variation in a sample of early/middle to late Holocene individuals from Peru. For this, eight linear measurements were recorded on the facial skeleton and cranial vault of 301 individuals coming from 19 archaeological sites located in four different biomes. Principal Component and Discriminant Analysis allowed exploring the skull shape and size variation. Complementarily, morphological affinity among the samples was studied by calculating Mahalanobis distances and Ward’s hierarchical cluster analysis. Additionally, the effects of biome, altitude, archaeological site, and chronology on morphology were evaluated through a MANOVA. The results showed that there are similarities between individuals of different chronology; the early/middle Holocene individuals show large morphological variation; and the most recent samples show differences in the length of the skull and nasal variables, but also in the alveolar subcomponent, which in turn are associated with the diversity in altitude and archaeological site of origin. The results are discussed in the context of previous works that studied samples from Peru, as well as some other areas of South America. In this way we hope to contribute to the current debates about the population of South America, especially in relation to local adaptation.
Keywords: Morphological evolution, Ecological variation, Early to late Holocene, Central Andes.
Resumo: Estudo ecomorfológico da variação craniana em populações pré-históricas do Perú. O objetivo deste trabalho é analisar a variação craniométrica em uma amostra de indivíduos do Holoceno inicial/médio ao final do Peru. Para isto, foram registradas oito medidas lineares no esqueleto facial e abóbada craniana de 301 indivíduos procedentes de 19 sítios arqueológicos, os quais estão localizados em quatro biomas. Explorou-sea variação na forma e tamanho do crânio entre indivíduos de diferentes biomas e sítios arqueológicos por meio de uma Análise de Componentes Principais e Análise Discriminante. Além disso, a afinidade morfológica entre as amostras foi avaliada pelo cálculo de distâncias de Mahalanobis e uma análise de cluster hierárquica de Ward. Ademais, estimou-se o efeito do bioma, sítio arqueológico, altitude e cronologia para explicar o padrão de variação morfológica observado através de MANOVA. Os resultados mostraram que existem semelhanças entre indivíduos de diferentes cronologias, os indivíduos do Holoceno inicial/médio apresentam grande variação morfológica, as amostras mais recentes mostram diferenças no comprimento do crânio, variáveis que descrevem a abertura nasal, mas também o subcomponente alveolar, que, por sua vez, está associada à diversidade de altitude e sítio arqueológico de origem. Os resultados são discutidos em relação a trabalhos prévios que estudaram amostras do Peru, e outras áreas da América do Sul. Desta maneira, espera-se contribuir para a discussão dos debates atuais sobre o povoamento da América do Sul, principalmente em torno das possíveis adaptações locais.
Palavras-chave: Evolução morfológica, Variação ecológica, Holoceno inicial a final, Andes Centrais.
Introducción
La variación morfológica inter-poblacional de grupos Americanos recientes ha sido descripta como sorprendentemente alta, incluso equivalente a la existente entre poblaciones de distintos continentes (Neves & Hubbe 2005; Sardi et al. 2005; González-José et al. 2008; Hubbe et al. 2015; von Cramon-Taubadel et al. 2017; Ponce de León et al. 2018; Ross & Ubelaker 2019). A pesar de que en las últimas décadas se han ofrecido múltiples explicaciones para comprender dicho patrón morfológico, recientemente se lo ha interpretado como resultado de un proceso de diversificación rápida debido a interacciones entre procesos aleatorios y fundamentalmente adaptaciones a diversos ecosistemas (Pérez et al. 2011; Menéndez 2015; de Azevedo et al. 2017). Sin embargo, aún queda por explicar detalladamente cómo se diferenciaron las poblaciones Sudamericanas en las distintas regiones ecológicas del continente, es decir, cuáles fueron los procesos evolutivos involucrados en la diferenciación poblacional de los grupos nativos americanos a lo largo del Holoceno. Para investigar esto resulta crucial llevar adelante estudios que permitan analizar exhaustivamente cuáles fueron los procesos de diversificación involucrados durante las primeras expansiones hacia las regiones que presentan ocupaciones, desde momentos tempranos hasta más recientes. Sólo unos pocos trabajos han estudiado los cambios morfológicos en individuos pertenecientes a una o más secuencia(s) diacrónica(s) que se encuentren circunscritas geográficamente. En términos generales, estos trabajos plantean una moderada a alta variación craneométrica entre las poblaciones Sudamericanas, especialmente en Argentina y Brasil, con acentuadas diferencias entre los individuos del Holoceno temprano/medio y tardío. En las poblaciones de Argentina, tales diferencias han sido principalmente entendidas como resultado de la plasticidad fenotípica debido fundamentalmente a la diversidad en la dieta entre las distintas poblaciones prehistóricas (Sardi et al. 2006; Paschetta et al. 2010; Pérez et al. 2011; Menéndez et al. 2014, 2015, 2017, 2019a). Por otro lado, entre las poblaciones de Brasil, tales diferencias han sido explicadas como producto de migraciones continentales sucesivas (Hubbe et al. 2014; von Cramon-Taubadel et al. 2017). Entre las poblaciones de Chile y Colombia, se ha descripto una gran afinidad y continuidad cronológica, lo cual ha sido interpretado en términos de aislamiento geográfico, pero complementado con flujo génico como producto de migraciones de grupos aledaños (Delgado 2015; Menéndez et al. 2015; Kuzminsky et al. 2017). Finalmente, un trabajo reciente ha interpretado las diferencias morfológicas entre individuos del Holoceno temprano/medio y tardío de Perú, como resultado de selección direccional asociada a diversidad climática y nutricional (Menéndez et al. 2019b).
El objetivo de este trabajo es evaluar cuáles fueron los procesos evolutivos involucrados durante la diversificación de las poblaciones prehistóricas de Perú. Particularmente, se analizará la variación que presentan un conjunto de individuos provenientes de distintos biomas, para inferir cuáles procesos evolutivos contribuyeron a su diferenciación. Perú constituye una de las áreas más indicadas para evaluar la influencia de la diversidad ecológica sobre la morfología esqueletal debido a que presenta una gran diversidad de biomas a lo largo y ancho de su extensión geográfica. Estudios previos realizados comparando cráneos de Perú han descripto diferentes tendencias en los datos y ofrecido múltiples interpretaciones: A) Ausencia de un patrón morfológico diferencial entre muestras de la costa y de tierras altas, como resultado del limitado flujo génico y la relevancia de la deriva génica, por lo que el principal eje de variación morfológica está dado por la proximidad geográfica entre las poblaciones, despreciando las similitudes entre regiones ecológicas (Stone et al. 2015; Butaric & Klocke 2018), B) Diferencias entre individuos de distintas áreas ecológicas, los de la costa caracterizados por cráneos cortos en sentido anteroposterior, los de tierras altas presentan cráneos alargados como resultado del aislamiento geográfico entre regiones ecológicas. Este patrón presenta dos alternativas: B1) Diferencias más pronunciadas en momentos tempranos y de menor magnitud hacia momentos más recientes, como producto del reciente incremento en el flujo génico entre dichas áreas, resultando en que los individuos recientes de tierras altas presentarían cráneos más cortos en comparación con los más antiguos de la misma región (Newman 1943; Ericksen 1962); B2) Diferencias entre tierras altas y tierras bajas se habrían incrementado en el tiempo como resultado de adaptaciones locales, según la diversidad en el clima, dieta y altitud de las distintas regiones ecológicas (Verano 1987; Ross et al. 2008). Si A fuese el escenario más plausible, entonces esperaríamos encontrar similitudes morfológicas entre individuos de distintos biomas. Por otro lado, si hay diferencias morfológicas entre individuos procedentes de distintos biomas, entonces B es la mejor explicación. Sin embargo, si las diferencias morfológicas se encuentran presentes entre los individuos más tempranos, y son bajas o nulas entre los individuos recientes, entonces B1 constituye la explicación más plausible. Por el contrario, si tales diferencias se incrementan con el tiempo, entonces B2 constituye la explicación adecuada para el presente estudio.
Para poner a prueba dichas hipótesis, en el presente trabajo, se evaluará la variación craneométrica en un conjunto de muestras de Perú en relación al sitio arqueológico de origen, bioma de procedencia, y cronología. La variación morfológica en forma y tamaño entre los grupos procedentes de distintos biomas y sitios arqueológicos será evaluada mediante Análisis de Componentes Principales (ACP), y Análisis Discriminante (AD). La cuantificación de dichas diferencias será evaluada mediante el cálculo de distancias Mahalanobis (D2) y un análisis jerárquico de clúster (Método de Ward). Finalmente, se evaluará si la variación morfológica se encuentra asociada a diferencias cronológicas y ecológicas mediante un Análisis Multivariado de la Varianza (MANOVA). Se discutirán los resultados obtenidos en relación a cuáles fueron los principales procesos evolutivos que contribuyeron a generar la variación morfológica de las poblaciones nativas de Perú, lo cual permitirá aportar reflexiones a los debates actuales sobre el poblamiento de Sudamérica.
Material y Métodos
Muestra
Se analizó un total de 301 cráneos de individuos adultos procedentes de 19 sitios arqueológicos de Perú (Fig.1, Tabla 1). Los datos fueron obtenidos de múltiples bases de datos, colectados anteriormente de manera independiente, y fueron recientemente reunidos para el presente trabajo: (1) medidas lineales calculadas a partir de una reconstrucción virtual generada mediante tomografías computadas del individuo 15.07 del sitio arqueológico Cuncaicha (Rademaker et al. 2014; Menéndez et al. 2019b), (2) 110 especímenes del sitio Yauyos, de la base de datos del Dr. William W. Howells (Howells 1973), y (3) una selección de 190 individuos, provenientes de 18 sitios arqueológicos, de la base de datos del Dr. Héctor Pucciarelli (Pucciarelli et al.2006, 2008, 2010, entre otros). Esta última se encuentra disponible online (http://www.repositorio.cenpat-conicet.gob.ar/handle/123456789/488), como Anexo del libro que fue publicado en honor a las contribuciones que realizó dicho investigador a la bioantropología latinoamericana (Paschetta et al. 2017). La misma fue utilizada en múltiples estudios que fueron realizados por otros investigadores comparando muestras en escala global o continental (Pérez et al. 2011; de Azevedo et al. 2012; Menéndez et al.2015, 2017).
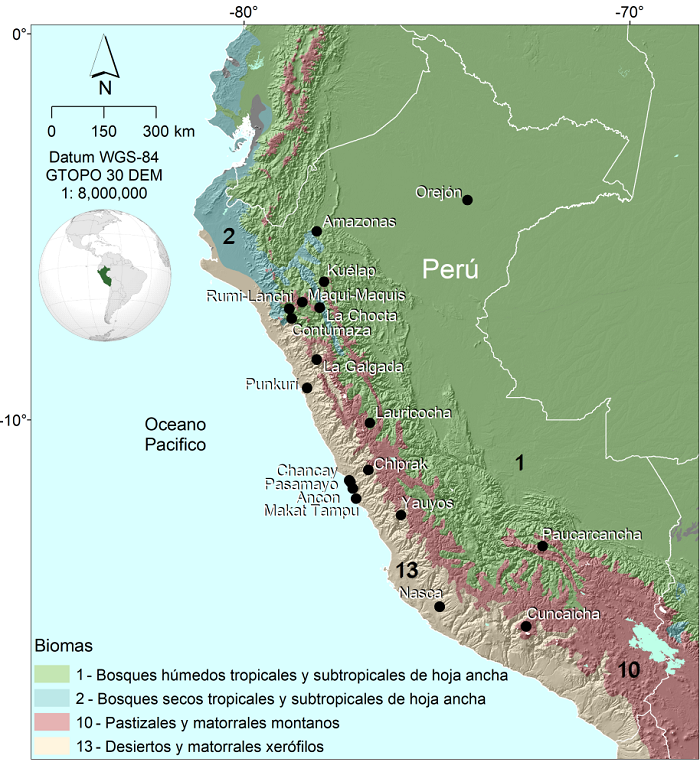
La totalidad de los individuos fue agrupada en base al (a) sitio arqueológico, (b) bioma de procedencia, y (c) cronología, tal como se describe en la Tabla 1. La información acerca de la ubicación de los sitios arqueológicos fue obtenida tanto de los catálogos de museos, como de publicaciones previas que analizaron dichas muestras. Luego, en base al trabajo de Olson y colaboradores (2001), los sitios fueron agrupados en los siguientes 4 biomas, de un total de 14 que se describen en dicho trabajo: (b1) Bioma 1: bosques húmedos tropicales y subtropicales de hoja ancha, (b2) Bioma 2: bosques secos tropicales y subtropicales de hoja ancha, (b10) Bioma 10: pastizales y matorrales montanos, y (b13) Bioma 13: desiertos y matorrales xerófilos. Dichas regiones fueron establecidas siguiendo criterios biogeográficos, según los cuales una ecoregión es definida como un área terrestre relativamente amplia que contiene un conjunto distintivo de especies y comunidades naturales (Olson et al. 2001). Finalmente, en relación a la cronología, sólo se dispone de fechados radiocarbónicos directos para las muestras de Lauricocha (Cardich 1964; Lindauer et al. 2015), Cuncaicha (Rademaker & Hodgins 2018), y La Galgada (Grieder et al. 1988; Posth et al. 2018). Los otros sitios presentan dataciones relativas asociadas al Holoceno tardío (Tabla 1; Howells 1973; Menéndez et al. 2019b). A los fines de los análisis, se utilizaron las fechas absolutas para los sitios con los que se cuenta con cronología precisa, y las medias de los rangos temporales para las muestras del Holoceno tardío, siguiendo a Walker et al. 2012 (4200 cal AP).
| Cronología | Bioma | Sitio Arqueológico | n (f/m) | Años calibrados AP (rango 95 %) | Referencia fechado radiocrabónico | Base de datos morfométricas |
| Holoceno temprano/ medio | Pastizales y matorrales montanos | Lauricocha (Lau) | 5 (1/4) | *8746-3450 cal AP | Cardich (1964); Lindauer et al. (2015) | Pucciarelli (2008) |
| Cuncaicha (Cun) | 1 (1/0) | *9133-8789 cal AP** | Rademaker & Hodgins (2018) | Menéndez et al. (2019b) | ||
| Holoceno tardío | Bosques húmedos tropicales y subtropicales de hoja ancha | Amazonas (Ama) | 8 (0/8) | Pucciarelli (2008) | ||
| Orejón (Ore) | 4 (0/4) | Pucciarelli (2008) | ||||
| Kuélap (Kue) | 8 (0/8) | 3000-500 cal AP | Pucciarelli (2008) | |||
| Rumi Lanchi (RLa) | 8 (0/8) | Pucciarelli (2008) | ||||
| Contumaza (Con) | 12 (0/12) | Pucciarelli (2008) | ||||
| Paucarcancha (Pau) | 10 (0/10) | Pucciarelli (2008) | ||||
| Bosques secos tropicales y subtropicales de hoja ancha | La Chocta (Cho) | 6 (0/6) | 3000-500 cal AP | Pucciarelli (2008) | ||
| Pastizales y matorrales montanos | Maqui-Maquis (MMa) | 10 (0/10) | Pucciarelli (2008) | |||
| Yauyos (Yau) | 110 (55/55) | 3000-500 cal AP | Howells (1973, 1989) | |||
| Desiertos y matorrales xerófilos | Nasca (Nas) | 26 (0/26) | Pucciarelli (2008) | |||
| Chiprak (Chp) | 3 (0/3) | Pucciarelli (2008) | ||||
| MakatTampu (MTa) | 20 (0/20) | 3000-500 cal AP | Pucciarelli (2008) | |||
| Chancay (Cha) | 15 (0/15) | Pucciarelli (2008) | ||||
| Punkuri (Pun) | 6 (0/6) | Pucciarelli (2008) | ||||
| Pasamayo (Pas) | 20 (9/11) | Pucciarelli (2008) | ||||
| Ancón (Anc) | 25 (0/25) | Pucciarelli (2008) | ||||
| La Galgada (Gal) | 4 (0/4) | 3000-500 cal AP | Grieder et al. (1988); Posth et al.(2018) | Pucciarelli (2008) | ||
| TOTAL | 301 |
Análisis Preliminares
Error Inter-observador
A pesar de las discrepancias que pudieran generarse como producto de combinar bases de datos colectadas por distintos investigadores, en este trabajo se priorizó el incremento de la cantidad de especímenes de Perú. El propósito fue poder representar de una mejor manera la variación morfológica presente en los distintos biomas aquí analizados. Particularmente, la inclusión de la muestra de Yauyos registrada por Howells, permite incrementar la cantidad de individuos procedentes del bioma “pastizales y matorrales montanos”. Por esta razón, en primer lugar, se seleccionaron meticulosamente medidas lineales que fueran equivalentes, según los criterios especificados por Howells (1973, 1989) y por Pucciarelli (2008) (Tabla 2). Sin embargo, dado que las mismas medidas pueden diferir cuando son registradas por distintos investigadores, se procedió a evaluar el error inter-observador durante su registro. Para esto, se seleccionaron 8 individuos procedentes de distintos continentes, que fueron medidos por ambos investigadores. Se calculó el Índice de Correlación Intraclase (ICC) para cada una de las 8 medidas seleccionadas para llevar adelante los siguientes análisis del presente trabajo (Shrout & Fleiss 1979; Menéndez 2017). Los resultados muestran que en cada caso, el ICC es mayor a 0,82, valor que ha sido considerado por Landis & Koch (1977) como casi perfecto, por lo que decidimos continuar con los análisis principales del trabajo.
| Pucciarelli | Howells | |||||
| Abr. | Nombre | Definición | Abr. | Nombre | Definición | |
| Neurocraneo | LNM | Longitud Neurocraneana Media | Bregma (Br) - Lambda (Lb). | PAC | Cuerda Bregma-Lambda o Cuerda Parietal (Bregma-Lambda chord; Parietal chord) | Distancia directa del Bregma al Lambda (The external chord, or direct distance from Bregma to Lambda, taken in the midplane and at the external surface). |
| ANP | Ancho Neurocraneano Posterior | Asterion (As) - Asterion (As). | ASB | Ancho Biasterion (Biasterionic Breadth) | Distancia directa del Asterion derecho al Asterion izquierdo (Direct measurement from one Asterion to the other). | |
| Esqueleto Facial | HF | Altura Facial | Nasion (Na) - Prostion (Pr). | NPH | Altura Nasion-Prostion (Nasion-Prosthion Height) | Distancia del Nasion al Prostion (Upper facial height from Nasion to prosthion). |
| AO | Ancho Óptico | Dacrion (Da) - Ectoconquio (Ek). | OBB | Ancho Orbital (Orbit Breadth) | Distancia del Ectoconquio al Dacrion (Breadth from Ectoconchion to Dacryon) | |
| HO | Altura Óptica | Supraobital (So) - Infraorbital (Io). | OBH | Altura Orbital (Orbit Height) | Distancia entre el borde superior e inferior de la órbita (The height between the upper and lower borders of the orbit, perpendicular to the long axis of the orbit and bisecting it). | |
| AR | Ancho Respiratorio | Alar izquierdo (Al) - Alar derecho (Al). | NLB | Ancho Nasal (Nasal Breadth) | Distancia máxima en los bordes anteriores de la apertura nasal (The distance between the anterior edges of the nasal aperture at its widest extent). | |
| HR | Altura Respiratoria | Nasion (Na) - Subnasal (Sn). | NLH | Altura Nasal (Nasal Heigth) | Distancia del Nasion al borde inferior de la apertura nasal (The average height from Nasion to the lowest point on the border of the nasal aperture on either side). | |
| AA | Ancho Alveolar | Ectomolar izquierdo (Em) -Ectomolar derecho (Em). Coincide con la mayor apertura del calibre. | MAB | Ancho del Paladar (Palate Breadth) | Apertura máxima entre los bordes alveolares (The greatest breadth across the alveolar borders, wherever found, perpendicular to the median plane). | |
Estandarización del sexo
Respecto a la composición de las muestras, en general los individuos no se encuentran balanceados por sexo (Tabla 1). Mientras que los individuos del Holoceno tardío de la base de datos de Pucciarelli son en su mayoría masculinos (excepto la muestra de Pasamayo que presenta 11 individuos determinados como masculinos, y nueve como femeninos), los individuos de los sitios del Holoceno temprano/medio son tanto masculinos como femeninos (Pucciarelli et al. 2006). Por el contrario, la muestra de Yauyos, sí se encuentra balanceada por sexo, la mitad de los individuos son masculinos y la otra mitad son femeninos (Howells 1973). Finalmente, el individuo 15.07 de Cuncaicha es un adulto joven de sexo femenino (Francken et al. 2018). Se realizó un análisis de la varianza para evaluar si existen diferencias en forma y tamaño debida al sexo, y las mismas resultaron significativas (<0,01). Dadas estas discrepancias, se realizó una estandarización del sexo para evitar introducir en los análisis posteriores variación debida al efecto del dimorfismo sexual. Siguiendo a Ackermann & Cheverud (2002), esto fue realizado mediante la adición del resto obtenido a partir de la diferencia media entre individuos masculinos y femeninos, a cada individuo de sexo femenino. Dichos cálculos se realizaron por separado para cada población de la muestra. Los siguientes análisis fueron realizados con las variables estandarizadas.
Variables morfométricas
La variación en la forma del cráneo fue estudiada mediante la selección de un conjunto de distancias lineales que constituyen algunas de las distancias tradicionalmente utilizadas en estudios antropológicos (Fig. 2). Las mismas incluyen medidas que describen tanto la forma del esqueleto facial como de la bóveda craneana. Tal como se mencionó anteriormente, dichas medidas fueron definidas y registradas independientemente por Héctor M. Pucciarelli y por William W. Howells.
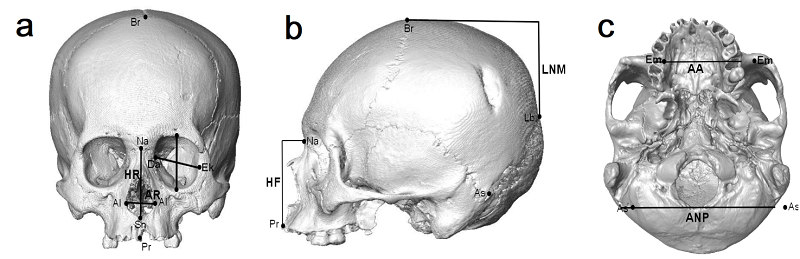
Referencias según la nomenclatura de Pucciarelli: HF [Altura Facial: Nasion (Na) – Prostion (Pr)]; LNM [Longitud Neurocraneana media: Bregma (Br) – Lambda (Lb)]; ANP [Ancho Neurocraneano Posterior: Asterion (As) – Asterion (As)]; AO [Ancho Óptico: Dacrion (Da) – Ectoconquion (Ek)]; HO [Altura Óptica: Supraorbital (So) – Infraorbital (Io)]; AR [Ancho Respiratorio: Alar (Al) – Alar (Al)]; HR [Altura Respiratoria: Nasion (Na) - Subnasal (Sn)]; AA [Ancho Alveolar: Ectomolar (Em) – Ectomolar (Em)].
La base de datos de Pucciarelli contiene un total de 30 medidas lineales, las cuales fueron registradas siguiendo el método craneofuncional (Pucciarelli 2008; Sardi 2017). Por su parte, la base de datos de Howells presenta un total de 82 medidas lineales que describen el cráneo de manera exhaustiva (Howells 1973). Sin embargo, dichos investigadores contaban con criterios diferentes en la definición y registro de medidas. Mientras que el propósito de Howells era definir la forma del cráneo de manera amplia mediante un conjunto de medidas distribuidas en todas las regiones del mismo (Howells 1973), Pucciarelli estaba interesado en medidas craneofuncionales que permitan dilucidar el efecto biológico y las múltiples causas de la variación humana (Pucciarelli 2008; Sardi 2017). Es decir, la base de datos de Pucciarelli presenta medidas que describen regiones más pequeñas pero con origen embriológico, contenido tisular, y funciones claramente definidas, mientas que la de Howells parte de las medidas convencionales y las modifica, agregando y quitando algunas, en base a su experiencia. Dados los distintos propósitos, desafortunadamente sólo existen pocas medidas comunes entre ambos investigadores (Tabla 2). Además, el estado de conservación del individuo de Cuncaicha no permite registrar muchas de las medidas definidas por dichos autores. De manera que, finalmente, siguiendo las definiciones de las medidas de ambos conjuntos de datos, y en base al estado de conservación del individuo de Cuncaicha, se seleccionaron un total de ocho distancias lineales para el presente estudio: dos correspondientes a la bóveda, y seis al esqueleto facial, dado que este último presenta mejores condiciones de conservación. Dentro del esqueleto facial, dos de las medidas describen el subcomponente óptico, dos el respiratorio, una el alveolar, y una la altura facial. Las medidas del individuo de Cuncaicha se registraron sobre el modelo de cráneo reconstruido virtualmente utilizando el programa Avizo (Menéndez et al. 2019b).
Se obtuvieron variables de forma a partir del cálculo de los radios de forma de Mosimann (Darroch & Mosimann 1985). Para esto, en primer lugar se calculó el tamaño, el cual fue estimado mediante la media geométrica para cada población de la muestra. Luego, se calculan las variables de forma a partir del cociente entre las variables originales de cada individuo y la media geométrica de todas las variables para dicho individuo (Menéndez & Pérez 2011). Los análisis posteriores fueron realizados a partir de las variables de forma y de tamaño de manera independiente.
Análisis estadísticos
La variación en forma entre las muestras provenientes de distintos biomas fue estudiada mediante análisis estadísticos exploratorios tales como el ACP y AD. Estos análisis fueron realizados para explorar la distribución de los especímenes de acuerdo a las similitudes y diferencias morfológicas entre sí. Los mismos permiten calcular ejes de variación independientes que posibilitan distinguir aquellas variables de forma que principalmente contribuyen a describir la variación presente en la muestra (ACP), así como aquellas que contribuyen a maximizar las diferencias entre los grupos en los que se clasifica a los individuos (AD) (Zar 2010). Se presenta la distribución de la variación a lo largo de los cuatro primeros componentes principales, y de los dos primeros ejes discriminantes. Para facilitar las interpretaciones, las muestras fueron agrupadas por sitio arqueológico (polígonos convexos, símbolos) y bioma (colores). Adicionalmente, para determinar cuáles variables principalmente contribuyen a generar dicho patrón de distribución, se presentan las cargas de las variables para cada uno de los componentes principales y de los ejes discriminantes. Se describe la variación en tamaño de los individuos según el sitio arqueológico y bioma de procedencia mediante el análisis de gráficos de caja. Los mismos muestran la mediana de la distribución, los cuartiles Q1 (25 % de los datos) y Q3 (75 % de los datos), valores máximo y mínimo (bigote de la caja), y valores atípicos (círculos por fuera de las cajas). Complementariamente, con el fin de evaluar cuantitativamente las similitudes y diferencias en forma entre las muestras provenientes de distintos sitios arqueológicos, se calcularon distancias generalizadas de Mahalanobis (D2), y se realizó un análisis de clúster jerárquico mediante el método de varianza mínima de Ward (Ward 1963). El cálculo de las D2 se basa tanto en la media y varianza de las variables predictoras, como en la matriz de covarianza de todas las variables. Son ampliamente utilizadas para analizar afinidades entre grupos debido a que no otorgan demasiado peso a variables que se encuentren correlacionadas entre sí. Además, presentan ventajas en estudios comparativos dado que toman en consideración la correlación del conjunto de datos en escala invariante (Mahalanobis 1930). El método de varianza mínima de Ward (Ward 1963) constituye un procedimiento que permite estimar grupos jerárquicos a partir de subconjuntos que contienen individuos que presentan similitudes entre sí. Se calcula un algoritmo recursivo que, en cada etapa, une los dos clústeres para los cuales se tenga el menor incremento en el valor total de la suma de los cuadrados de las diferencias, dentro de cada clúster, de cada individuo al centroide del clúster. Las distancias iniciales del clúster en el método de varianza mínima de Ward se definen como el cuadrado de la distancia euclidiana entre muestras. Estos análisis fueron realizados con el programa Past 3.15 (Hammer et al. 2001).
Finalmente, mediante un análisis multivariado de la varianza (MANOVA) se evaluó la existencia de diferencias estadísticas en la variación de la forma del cráneo (CP1, CP2, CP3, CP4) según el efecto del bioma, sitio arqueológico, altitud, y cronología. Adicionalmente, mediante un análisis univariado de la varianza (ANOVA), se evaluó la existencia de diferencias entre el tamaño del cráneo y el mismo conjunto de variables independientes. Con el fin de evaluar cuáles efectos están contribuyendo mayormente a los modelos obtenidos, se utilizaron el rastro de Pillai (MANOVA), y el test de Levene (ANOVA). El rastro de Pillai es un estadístico (P) que varía entre 0 y 1, de manera que los valores más altos indican que dicha variable contribuye en mayor medida al modelo en comparación a las otras. Si el valor de Pillai es cercano a 1, se debe rechazar la hipótesis nula de homogeneidad en la matriz de varianza-covarianza, mientras que si es cercano a 0 se considera que las matrices de varianza-covarianza no son diferentes (Pillai 1977). Respecto al test de Levene, si el valor p resultante es menor que el nivel de significancia (0,05), se rechaza la hipótesis nula de varianzas iguales y se concluye que existe una diferencia entre las varianzas de las muestras. Por lo tanto, en los resultados del MANOVA, se presentan los valores del rastro de Pillai (P), el F estimado a partir del mismo, y su valor significancia estadística (p). Los resultados del ANOVA se presentan mediante un estadístico (F) y su significancia asociada, la cual fue calculada a partir del test de Levene (p). Estos análisis fueron realizados en R (R Core Team 2013).
Resultados
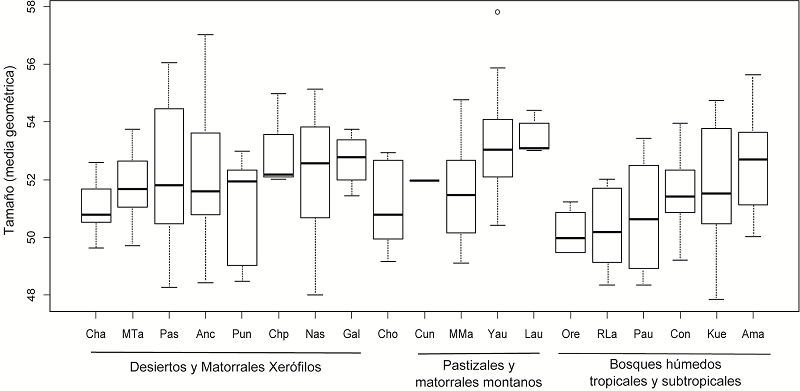
En primer lugar, se realizaron análisis exploratorios para evaluar la variación craneométrica en tamaño y forma que presentan los individuos de Perú mediante un gráfico de cajas, ACP y AD respectivamente. En la Figura 3 se exhibe el gráfico de caja que muestra la variación en tamaño por sitio arqueológico. Las muestras a su vez se encuentran agrupadas por bioma, los cuales está ordenados en sentido este-oeste. En general, se observa un grado alto de variación al interior de cada bioma. Sin embargo, se pueden describir algunas tendencias generales. Las muestras más tempranas (Cuncaicha, Lauricocha) presentan cráneos de tamaño moderado a alto. Si se consideran sólo las medianas, se puede observar que las muestras de “pastizales y matorrales montanos” presentan los cráneos más grandes del conjunto, tal es el caso para Yauyos y Lauricocha. Por otro lado, la muestra de Orejones presenta los cráneos más pequeños de todos los individuos aquí estudiados.
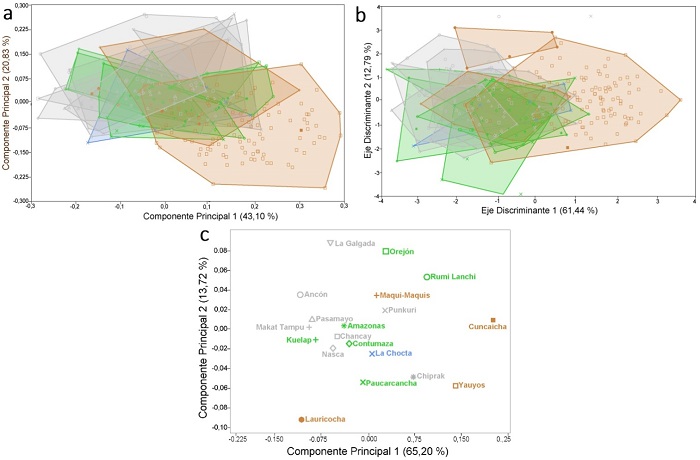
En la Figura 4a se muestra la variación morfológica a lo largo de los dos primeros componentes principales (CP), que juntos explican el 64 % de la variación total de la muestra (Tabla 3).Complementariamente, el ACP muestra que existe un alto grado de superposición en la variación en forma de las muestras analizadas. Sin embargo, a lo largo del CP1 se pueden visualizar algunas tendencias. Las muestras del bioma 10 “pastizales y matorrales montanos” se encuentran mayormente localizadas hacia el extremo positivo del CP1, mientas que las del bioma 13 “desiertos y matorrales xerófilos” se encuentran hacia el extremo negativo del CP1. Las muestras del bioma “bosques húmedos tropicales y subtropicales” y las de “bosques secos tropicales y subtropicales” se encuentran en el centro de la distribución. Este patrón se mantiene cuando se grafica el CP1/CP3, CP1/CP4, CP2/CP3, CP2/CP4, pero no con el CP3/CP4 donde la superposición de las muestras es aún mayor (resultados no mostrados). Las muestras tempranas no se distinguen de las recientes, el individuo de Cuncaicha se encuentra dentro del polígono convexo que agrupa a los individuos de Yauyos, mientras que las de Lauricocha se encuentran entre el centro de la distribución y el extremo negativo del CP1. Las variables que mayormente contribuyen a explicar dicha variación morfológica son la longitud neurocraneana media y altura respiratoria, aunque también el ancho neurocraneano posterior y la altura facial (Tabla 3). De manera adicional, en la Figura 4b, se presenta la variación a lo largo de los primeros ejes discriminantes, que juntos explican el 74 % de la variación total de la muestra. Las tendencias son similares, con los individuos del bioma “pastizales y matorrales montanos” mayormente localizados hacia el extremo positivo del primer eje discriminante, las de “desiertos y matorrales xerófilos” y “bosques húmedos tropicales y subtropicales” hacia el extremo negativo, y la de “bosques secos tropicales y subtropicales” en el centro. Las variables que mayormente contribuyen a dicha variación son la longitud neurocraneana media, la altura respiratoria, y el ancho neurocraneano posterior (Tabla 3). Complementariamente, el segundo eje discriminante distingue sutilmente la muestra de “bosques húmedos tropicales y subtropicales” hacia el extremo negativo, y la de “desiertos y matorrales xerófilos” hacia los valores positivos. Las variables que principalmente contribuyen a dicha variación son la altura facial, la altura óptica, y el ancho neurocraneano posterior (Tabla 3). Finalmente, la Figura 4c exhibe la variación de las medias de cada muestra a lo largo de los dos primeros componentes principales, que resumen un 78 % de la variación total. Los resultados son similares a los previos, las muestras de “pastizales y matorrales montanos” ubicadas hacia el extremo positivo del CP1, las de “desiertos y matorrales xerófilos” hacia el extremo negativo del CP1 y positivo del CP2, las de “bosques húmedos tropicales y subtropicales” en el centro y extremo positivo del CP2.
| CP 1 (43,1 %) | CP 2 (20,8 %) | CP 3 (14,4 %) | CP 4 (10,1 %) | ED 1 (61,4 %) | ED 2 (12,8 %) | |
| Longitud Neurocraneana Media (LNM) | 0,890 | -0,344 | 0,161 | 0,187 | 0,076 | -0,015 |
| Ancho Neurocraneano Posterior (ANP) | 0,287 | 0,926 | 0,057 | 0,181 | -0,022 | 0,020 |
| Altura Facial (HF) | -0,251 | -0,123 | 0,475 | 0,545 | 0,003 | 0,038 |
| Ancho Óptico (AO) | 0,005 | 0,006 | -0,025 | -0,236 | -0,001 | -0,009 |
| Altura Óptica (HA) | -0,093 | 0,008 | 0,131 | -0,172 | -0,008 | -0,015 |
| Ancho Respiratorio (AR) | 0,004 | -0,049 | -0,213 | -0,367 | 0,004 | -0,009 |
| Altura Respiratoria (HR) | -0,217 | -0,019 | 0,392 | 0,211 | -0,028 | 0,010 |
| Ancho Alveolar (AA) | -0,080 | -0,138 | -0,727 | 0,609 | 0,001 | 0,014 |
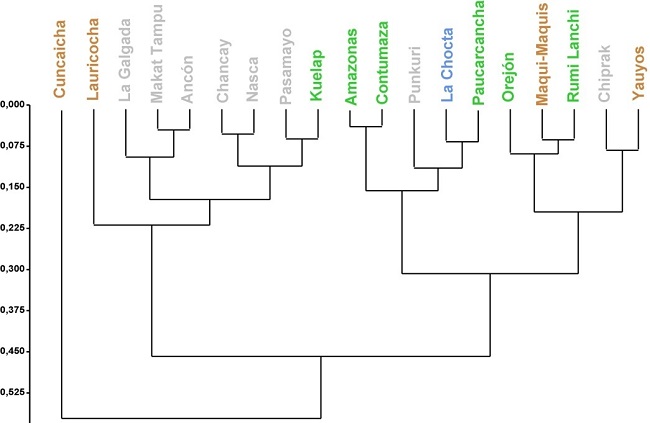
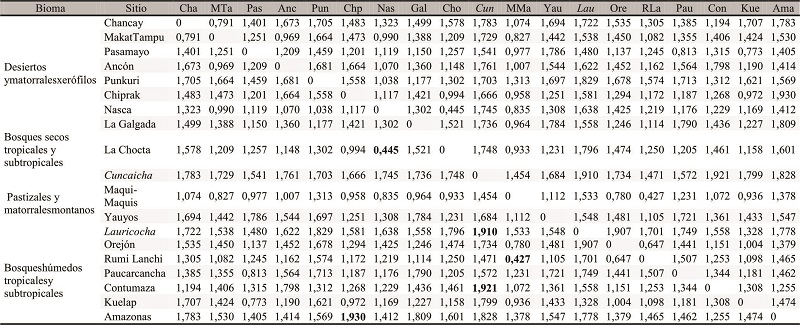
A continuación, mediante el cálculo de Distancias Mahalanobis y la estimación del análisis de clúster jerárquico de Ward se evaluó la magnitud de las distancias fenotípicas entre los grupos. De acuerdo con las primeras, las muestras que presentan mayores similitudes entre sí son La Chocta/Nasca, y Maqui-Maquis/Rumi Lanchi, mientras que las que más difieren entre sí son las de Lauricocha/Cuncaicha, Contumaza/Cuncaicha, y Chiprak/Amazonas (Tabla 4). Las similitudes entre Maqui-Maquis y Rumi Lanchi se pueden interpretar en términos de cercanía geográfica, pero las otras similitudes y diferencias son más difíciles de interpretar. Todos los pares de muestras pertenecen a distintos biomas (excepto las muestras tempranas de Lauricocha y Cuncaicha que ambas pertenecen a “pastizales y matorrales montanos”), por lo que el bioma no parecería jugar un rol en término de similitud o diferencia entre las muestras. Asimismo, cuando se analizan las muestras del Holoceno temprano/medio (Cuncaicha/Lauricocha), las mismas presentan grandes diferencias entre sí. A su vez, el individuo de Cuncaicha presenta mayores similitudes con los de Rumi Lanchi y Maqui-Maquis, mientras que los de Lauricocha con los de Kuelap y Pasamayo. El análisis de clúster jerárquico de Ward muestra, en principio que la muestra de Cuncaicha, la más antigua, difiere considerablemente del resto (Fig. 5). Luego, las muestras restantes se agrupan en dos grandes clústeres, uno que contiene muestras procedentes principalmente del bioma “desiertos y matorrales xerófilos”, y otro con la mayor parte de las muestras de “bosques húmedos tropicales y subtropicales” y la única de “pastizales y matorrales montanos” (Fig. 5). La muestra de Lauricocha, a pesar de que forma parte del clúster “desiertos y matorrales xerófilos”, parece diferenciarse considerablemente de las demás.
Finalmente, mediante MANOVA, se analizó el efecto de variables como el bioma, altitud, sitio arqueológico, y cronología para explicar su efecto sobre la variación craneométrica de la muestra. Los resultados muestran que la mayor parte de la variación en forma en el cráneo se puede interpretar como resultado de las diferencias en sitio arqueológico (P= 0,49) y altitud (P= 0,31), el bioma tendría un efecto muy bajo (P= 0,10). Estos 3 factores presentan contribuciones al modelo que son estadísticamente significativas (<0,01). La cronología presenta una contribución despreciable (P= 0,02) y no significativa (p= 0,18), es decir no existen diferencias según período cronológico (Tabla 5). Es decir, la hipótesis nula de homogeneidad de la matriz de varianza-covarianza cuando se comparan individuos procedentes de distintos biomas, sitios de distinta altitud, y/o sitios arqueológicos, debe ser rechazada, dado que dichas muestras difieren de manera considerable entre sí. Tales resultados parecen coincidir con los de las distancias Mahalanobis y el de clúster jerárquico de Ward. Finalmente, cuando se comparan las diferencias en tamaño del cráneo, existen diferencias estadísticamente significativas según la altitud y sitio arqueológico de procedencia. Sin embargo, cronología y bioma tienen un efecto despreciable en el modelo (Tabla 5). Es decir, forma y tamaño parecen variar como resultado de los mismos factores ecológicos: altitud, y sitio arqueológico de procedencia.
| Forma del Cráneo | Tamaño del Cráneo | |
| Sitio arqueológico | P= 0,49 F= 2,65 p<0,01 | F= 3,35 p= 0,01 |
| Altitud | P=0,31 F=31,32 p<0,01 | F= 37,5 p= 0,01 |
| Bioma | P= 0,10 F= 8,22 p<0,01 | F= 9,92 p= 0,15 |
| Cronología | P= 0,02 F= 1,56 p= 0,18 | F= 0,22 p= 0,69 |
Discusión
Los análisis realizados en el presente trabajo estuvieron dirigidos a evaluar la existencia de diferencias morfológicas entre individuos provenientes de distintos biomas y periodos cronológicos de Perú. Perú es un área con una gran diversidad de biomas, lo que permite discutir la interacción entre la diversidad ecológica y los procesos evolutivos involucrados durante la diversificación de las poblaciones humanas en los Andes Centrales. Los resultados aquí obtenidos apoyan estudios craneométricos previos que afirman que las poblaciones humanas de los Andes presentan continuidad biológica (Delgado 2015; Kuzminsky et al. 2017). Es decir, contradicen argumentos previos donde se ha planteado que existen grandes diferencias entre muestras tempranas y recientes de Sudamérica (Neves & Pucciarelli 1991; Neves & Hubbe 2005; von Cramon-Taubadel et al. 2017, entre otros). Los resultados del Análisis de Componentes Principales, Analisis Discriminante y la comparación de las distancias de Mahalanobis muestran que los individuos tempranos presentan similitudes con los tardíos. Particularmente, el individuo de Cuncaicha presenta similitudes con los de Yauyos, Rumi Lanchi y Maqui-Maquis, mientras los de Lauricocha con Pasamayo y Kuélap. De estos últimos, sólo los sitios de Cuncaicha, Maqui-Maquis, y Yauyos se encuentran ubicados en el mismo bioma (“pastizales y matorrales montanos”), por lo que las similitudes pueden ser interpretadas como resultado de continuidad biológica y/o adaptaciones a un mismo ambiente. Además, los resultados del MANOVA muestran que no existen grandes diferencias entre los individuos del Holoceno temprano/medio y aquellos del Holoceno tardío, a pesar que el número de individuos de momentos tempranos aquí analizados sea muy bajo.
A pesar de tales limitaciones debidas al bajo número de individuos del Holoceno temprano/medio en la muestra analizada, el rango de variación de los mismos parece ser amplio (a lo largo del primer componente principal, y las distancias Mahalanobis), lo cual brinda soporte a la idea de que la variación morfológica entre los individuos tempranos haya sido alta (Pucciarelli et al. 2003; Sardi et al.2004, 2005; González-José et al. 2008). Los resultados aquí obtenidos confirman que este sería un patrón presente no sólo cuando se comparan individuos de áreas lejanas sino también de una misma región, como es el caso de los Andes centrales. Sin embargo, debe tenerse en cuenta que esto puede ser producto de un artefacto metodológico, donde si bien la variable cronológica fue cuantificada de manera continua, solo 4 de las 19 muestras presentan fechados radiocarbónicos directos. Para las otras muestras se utilizó una media que equivale al rango cronológico del Holoceno tardío. Por esta razón, esta variable se expresa de manera constante para dichos casos, en lugar de manera ordinal. Esta decisión metodológica puede estar afectando los mencionados resultados del MANOVA. A pesar de esto, es sobresaliente el hecho de que las mayores diferencias se encuentren entre las muestras del Holoceno temprano, y en segundo lugar, entre muestras del Holoceno tardío entre sí, mientras que las diferencias entre muestras del Holoceno temprano/medio y tardío no son considerables. Este resultado brinda apoyo al modelo que plantea que la evolución de las poblaciones humanas de Sudamérica es el resultado de diversos procesos evolutivos rápidos ocurridos in situ y no de distintas oleadas migratorias (Pérez & Monteiro 2009; Pérez et al. 2009; Menéndez et al. 2014; de Azevedo et al. 2017).
Asimismo, el ACP y el cálculo de las distancias Mahalanobis confirmaron grandes similitudes morfológicas entre las muestras del Holoceno tardío. Las mismas pueden explicarse como resultado de migraciones frecuentes entre individuos pertenecientes a poblaciones que se encontraban en distintas áreas. Esto brinda apoyo a algunos trabajos previos que describieron la existencia de una varianza baja entre poblaciones de los Andes, fundamentalmente debida a aislamiento geográfico, especialmente entre muestras de los andes/amazonia, y/o costa/sierra (Hrdlička 1911, 1914; Newman 1943; Tarazona-Santos et al. 2001; Ross et al. 2008). Dicho patrón puede ser resultado de dinámicas poblacionales con matrimonios preferenciales basadas en la exogamia y/o exoginia, tal como ha sido reportado recientemente para poblaciones de los Andes, Amazonia, y la costa Pacífica, basados en una combinación de datos genéticos y lingüísticos (Arias et al.2017, 2018; Barbieri et al. 2019). La presencia de sociedades de gran escala, tanto en los Andes como en la costa, puede haber promovido el flujo génico entre individuos, así como también combinado poblaciones que carecían de contacto previo o que era muy esporádico.
Los resultados aquí obtenidos confirman que gran parte la variación morfológica de las poblaciones de los Andes Centrales puede ser explicada como producto de adaptaciones a las diversas condiciones ambientales características del oeste de Sudamérica (Delgado 2017; Menéndez et al. 2019b). A partir de la simplificación de modelos previos, se puso a prueba si existe un patrón morfológico diferencial entre muestras de la costa y tierras altas (A-Ausencia de patrón ecomorfológico; B-Presencia de patrón ecomorfológico), y luego se exploró si el mismo habría estado presente desde momentos tempranos o si se habría establecido hacia momentos más recientes (B1-Patron morfológico antiguo; B2-Patron morfológico reciente). El resultado más relevante muestra que a pesar de que las diferencias morfológicas entre los individuos procedentes de distintos biomas son de baja magnitud, las diferencias altitudinales y el sitio arqueológico de procedencia habrían tenido un rol relevante durante la diferenciación morfológica de estas poblaciones. Es decir, nuestros resultados brindan soporte al Modelo B, describiendo la presencia de un patrón de variación morfológica en el cráneo asociado a la variación ecológica del área. Esto contradice estudios previos donde no se encontraron diferencias morfológicas entre individuos de tierras altas/tierras bajas y/o sierra/costa (Stone et al. 2015;Butaric & Klocke 2018). Asimismo, las similitudes morfológicas entre el individuo de Cuncaicha y los de Yauyos apoyan la hipótesis de que el patrón morfológico de adaptación ecológica se podría haber establecido desde momentos tempranos, ya que dichas muestras proceden del mismo bioma (“pastizales y matorrales montanos”). Este resultado podría confirmar la existencia de una temprana divergencia poblacional en los Andes Centrales (Harris et al. 2018). Desafortunadamente, el escaso número de muestras analizadas no permite interpretar esto de manera conclusiva. Por la misma razón, resulta difícil discernir si dicho patrón se mantuvo, incrementó, o disminuyó con el transcurso de las generaciones. Por lo que los dos modelos, el de diferencias más pronunciadas en momentos tempranos (Newman 1943; Ericksen 1962), o el incremento de diferencias morfológicas en el tiempo (Verano 1987; Ross et al. 2008) resultan igualmente plausibles.
De especial interés son los resultados obtenidos en relación al patrón de variación en el cráneo según los sitios arqueológicos, altitud, y bioma de procedencia. En relación al tamaño, existen diferencias estadísticamente significativas, según el sitio arqueológico y la altitud. Los individuos de “pastizales y matorrales montanos” (bioma de mayor altitud) presentan los cráneos más grandes del conjunto. En cuanto a los resultados de forma, la principal variación se produce en la longitud neucraneana media y altura respiratoria, aunque también el ancho neurocraneano posterior, altura facial, ancho respiratorio, y ancho alveolar contribuyen de manera relevante a lo largo de los primeros 4 componentes principales y 2 ejes discriminantes. Particularmente, los individuos de “pastizales y matorrales montanos” presentarian cráneos más alargados en sentido anteroposterior, esqueletos faciales más cortos, y aperturas nasales más anchas, en comparación a aquellos de los ‘bosques húmedos”, “bosques secos”, y “desiertos y matorrales xerófilos”, es decir a ambos lados de las estribaciones de los Andes. Estos resultados coinciden con trabajos previos que han planteado que los individuos de la costa se caracterizan por presentar cráneos cortos en sentido anteroposterior, mientras que los de tierras altas presentan cráneos alargados, diferencias que son interpretadas como resultado del aislamiento geográfico entre regiones ecológicas (Verano 1987; Ross et al. 2008). Asimismo, estudios previos plantean que factores no-aleatorios podrian haber tenido un rol relevante en la diversificación de las poblaciones de Perú (Menéndez et al. 2019b). Lo novedoso de este trabajo es que no sólo existen diferencias según la longitud de la bóveda craneana, lo cual puede ser interpretado como producto de la historia poblacional, sino que también las variables que describen las estructuras nasales, forma del esqueleto facial y del sucomponente alveolar presentan una amplia variación. De manera que tales diferencias morfológicas pueden resultar de adaptaciones, especialmente cuando se considera la forma de la nariz en relación a la presión ambiental del oxígeno y/o el rango de temperaturas anuales en los distintos biomas (Evteev et al. 2014; Butaric & Klocke 2018).
Con el fin de abordar de manera holística la complejidad de dimensiones imbuidas en el proceso de diversificación de las poblaciones en Sudamérica, estudios futuros que contribuyan a enriquecer los resultados aquí presentados deben considerar una mayor cantidad de variables en sus modelos, optimizar la identificación de los cambios morfológicos, y en la medida de lo posible, incrementar el número de individuos del Holoceno temprano/medio.
Conclusiones
El objetivo de este trabajo ha sido analizar la variación craneométrica en una muestra de individuos de Perú procedentes de 4 biomas y con asignaciones cronológicas que abarcan del Holoceno temprano al tardío, en relación al sitio arqueológico de origen, bioma de procedencia, altitud, así como también la cronología. Los resultados aquí presentados presentan limitaciones debidas fundamentalmente a los escasos individuos del Holoceno temprano/medio, y la baja cantidad de variables craneométricas, que además constituyen medidas lineales en lugar de puntos en 3D. Sin embargo, se pueden describir algunas tendencias: 1) las poblaciones de Perú presentan una continuidad biológica desde el Holoceno temprano/medio al tardío, 2) las poblaciones del Holoceno temprano/medio presentan considerable variación, 3) las poblaciones más recientes no presentan una amplia variación, 4) la variación morfológica no difiere considerablemente a lo largo del tiempo, 5) los principales cambios morfológicos se presentan en la longitud del cráneo, pero también en relación a variables que describen los subcomponentes nasal y alveolar, y 6) los principales factores de variación se encuentran asociados a la diversidad del sitio arqueológico, altitud, y en menor medida bioma de procedencia. Es decir, la diversificación morfológica entre poblaciones de Perú habría comenzado tempranamente. Estos resultados contribuyen a discutir la dinámica poblacional de Perú desde el momento de arribo de las primeras poblaciones hasta momentos históricos. Estudios futuros que incluyan una mayor cantidad de variables independientes, un mayor número de individuos, y variables morfológicas que permitan describir el cráneo de manera más completa, permitirán esclarecer con mayor detalle los procesos evolutivos que generaron dicha diversificación morfológica.
Agradecimientos
Una versión preliminar de este trabajo fue presentada en la sesión “El poblamiento de América y la Antropología Biológica. El desafío de la Interdisciplina”, coordinada por Josefina Motti y Rolando Gonzalez-José, como parte del 9° Simposio Internacional “El Hombre Temprano en América: La gente y sus lugares”, realizado en la ciudad de Necochea entre el 27 y el 30 de noviembre de 2018. Agradecemos a los organizadores por la invitación para presentar allí los resultados preliminares de este trabajo, y a los editores de este volumen especial, por invitarnos a publicarlo: Josefina Motti, Celeste Weitzel, Natalia Mazzia, Damián Bozzutto, Darío Hermo y Laura Marchionni. El trabajo de campo del sitio Cuncaicha fue realizado gracias a financiamiento otorgado al Dr. Rademaker por parte de la Fundacion Alexander von Humboldt, la Pontificia Universidad Catolica de Lima, Max Planck Institute for the Science of Human History, DFG (FOR 2237, INST 37/706) y Northern Illinois University. La presentación del trabajo en el congreso y escritura de este manuscrito fue posible gracias al apoyo económico brindado por el Konrad Lorenz Institute. Finalmente, agradecemos a Mark Hubbe y un(a) revisor(a) anónimo/a por los enriquecedores comentarios que permitieron mejorar una versión previa de este trabajo.
Referencias
Ackermann, R.R. & Cheverud, J.M. (2002) “Discerning evolutionary processes in patterns of tamarin (Genus Saguinus) craniofacial variation.” American Journal of Physical Anthropology 117(3), pp. 260-271.
Arias, L., Barbieri, C., Barreto, G., Stoneking, M. & Pakendorf, B. (2017) “High-resolution mitochondrial DNA analysis sheds light on human diversity, cultural interactions, and population mobility in northwestern Amazonia.” American Journal of Physical Anthropology 165(9), pp. 1-18.
Arias, L., Schroder, R., Hubner, A., Barreto, G., Stoneking, M. & Pakendorf, B. (2018) “Cultural innovations influence patterns of genetic diversity in northwestern Amazonia.” Molecular Biology and Evolution 35(11), pp. 2719-2735.
Barbieri, C., Barquera, R., Arias, L., Sandoval, J.R., Acosta, O., Zurita, C., Aguilar-Campos, A., Tito-Álvarez, A.M., Serrano-Osuna, R., Gray, R. Mafessoni, F., Heggarty, P., Shimizu, K.K., Fujita, R., Stoneking, M, Pugach I, Fehren-Schmitz, L. (2019) “The current genomic landscape of western South America: Andes, Amazonia, and Pacific Coast”. Molecular Biology and Evolution 36(12), 2698-2713.
Butaric, L.N. & Klocke, R.P. (2018) “Nasal variation in relation to high‐altitude adaptations among Tibetans and Andeans”. American Journal of Human Biology 30(3), pp. e23104
Cardich, A. (1964) Lauricocha: fundamentos para una prehistoria de los Andes Centrales. Buenos Aires, Centro Argentino de Estudios Prehistóricos.
Darroch, J.N. & Mosimann, J.E. (1985) “Canonical and principal components of shape.” Biometrika 72(2), pp. 241–252.
de Azevedo, S., Quinto-Sanchez, M., Paschetta, C., & Gonzalez-José, R. (2017) “The first human settlement of the New World: A closer look at craniofacial variation and evolution of early and late Holocene Native American groups.” Quaternary International 431(B), pp. 152-167.
de Azevedo, S., Pucciarelli, H.M., Lanata, J.L. & González-José, R. (2012) “Identificando señales de evolución no estocástica en la morfología craneofacial de poblaciones humanas modernas.” Revista Argentina de Antropología Biológica 14(1), pp. 113-129.
Delgado, M.E. (2015)“Variación dental y craneofacial en el norte de los Andes durante el Pleistoceno y el Holoceno. Su relevancia para la discusión de la colonización temprana de Sudamérica”, Tesis doctoral inédita, La Plata, Universidad Nacional de La Plata, 375 pp.
Delgado, M.E. (2017) “Holocene population history of the Sabana de Bogotá region, northern South America: An assessment of the craniofacial shape variation.” American Journal of Physical Anthropology 162(2), pp. 350-369.
Ericksen, M.F. (1962) “Undeformed pre-Columbian crania from the north sierra of Peru.” American Journal of Physical Anthropology 20(2), pp. 209-222.
Evteev, A., Cardini, A.L., Morozova, I., & O'Higgins, P. (2014) “Extreme climate, rather than population history, explains mid‐facial morphology of Northern Asians.” American Journal of Physical Anthropology 153(3), pp. 449-462.
Francken, M., Beier, J., Reyes-Centeno, H., Harvati, K. & Rademaker, K. (2018) “The human skeletal remains from Cuncaicharockshelter, Peru.” En: Harvati, K., Jaeger, G., Reyes-Centeno, H. (eds.) New Perspectives on the Peopling of the Americas, Kerns Verlag, Tuebingen, pp. 125-152.
Gonzalez-José, R., Bortolini, M.C., Santos, F. & Bonatto, S.L. (2008) “The peopling of America: Craniofacial shape variation on a continental scale and its interpretation from an interdisciplinary view.” American Journal of Physical Anthropology 137(2), pp. 175-187.
Grieder, T., Bueno Mendoza, A., Earle Smith, C. & Malina, R.M. (1988) La Galgada, Peru. A preceramic culture in transition. Austin, University of Texas Press.
Hammer, Ø., Harper, D.A.T. & Ryan, P.D. (2001) “PAST: Paleontological statistics software package for education and data analysis”. Palaeontologia Electronica 4(1), [en línea]. Disponible en: http://palaeoelectronica.org/2001_1/past/issue1_01.htm
Harris, D.N., Song, W., Shetty, A.C., Levano, K.S., Cáceres, O., Padilla, C., Borda, V., Tarazona, D., Trujillo, O., Sánchez, C., Kesslera, M. D., Galarza, M., Capristano, S., Montejo, H., Flores-Villanueva, P. O., Tarazona-Santose, E., O’Connora,T.D. & Guio, H. (2018) “Evolutionary genomic dynamics of Peruvians before, during, and after the Inca Empire”. Proceedings of the National Academy of Sciences 115(28), pp. e6526-e6535.
Howells, W.W. (1973) Cranial variation in man: a study by multivariate analysis of patterns of difference among recent human populations, Cambridge, Harvard University Press.
Howells, W.W. (1989) Skull shapes and the map: craniometric analyses in the dispersion of modern Homo, Harvard University Press.
Hubbe, M., Okumura, M., Bernardo, D.V. & Neves, W.A. (2014) “Cranial morphological diversity of early, middle, and late Holocene brazilian groups: implications for human dispersion in Brazil.” American Journal of Physical Anthropology 155(4), pp. 546-558.
Hubbe, M., Strauss, A., Hubbe, A. & Neves, W.A. (2015) “Early south americans cranial morphological variation and the origin of American biological diversity.” PloS ONE 10(10), pp. e0138090.
Hrdlička, A. (1911) Some Results of Recent Anthropological Exploration in Peru. Smithsonian Miscellaneous Collections Washington, Smithsonian Institution 56(16).
Hrdlička, A. (1914) “Anthropological work in Peru in 1913, with notes on the pathology of the ancient Peruvians”, Smithsonian Miscellaneous Collections 61(18), pp. 1-69.
Kuzminsky, S.C., Coonerty, N.& Fehren-Schmitz, L. (2017) “A reassessment of human cranial indices through the Holocene and their implications for the peopling of South America.” Journal of Archaeological Science: Reports 11, pp. 709-716.
Landis, J.R. & Koch, G.G. (1977) “The measurement of observer agreement for categorical data.” Biometrics 33(1), pp. 159-174.
Lindauer, S., Tomasto-Cagigao, E.& Fehren Schmitz, L. (2015) “The skeletons of Lauricocha: New data on old bones.” Journal of Archaeological Science: Reports 4, pp. 387-394.
Mahalanobis, P.C. (1930) “On tests and measures of group divergence”. Journal and Proceedings of Asiatic Society of Bengal New series 26, pp. 541-588.
Menéndez, L.P. (2017) “Comparing methods to assess intraobserver measurement error of 3D craniofacial landmarks using geometric morphometrics through a digitizer arm.” Journal of Forensic Sciences 62(3), pp. 741-746.
Menéndez, L.P. & Pérez, S.I. (2011) “Información morfométrica en estudios Interpoblacionales: una comparación de técnicas basadas en proporciones y coordenadas Procrustes.” Revista Argentina de Antropología Biológica 13(1), pp. 105-111.
Menéndez, L.P., Bernal, V., Novellino, P. & Pérez, S.I. (2014) “Effect of bite force and diet composition on craniofacial diversification of southern south American human populations.” American Journal of Physical Anthropology 155(1), pp. 114-127.
Menéndez, L.P., Pérez, S.I., Pucciarelli, H.M., Bonomo, M., Messineo, P.G., Gonzalez, M.E. & Politis, G.G. (2015) “Early Holocene human remains from the argentinean pampas: cranial variation in south America and the American peopling.” PaleoAmerica 1(3), pp. 251-265.
Menéndez, L.P., Bonomo, M., Messineo, P.G., Gonzalez, M.E., Politis, G.G. & Pérez, S.I. (2017) “Craniofacial variation of early americans from argentinean pampas: Implications for the settlement of the New World.” En: Paschetta, C., González-José, R. &Lanata, J.L. (eds.) De cómo cruzar fronteras en la ciencia. Homenaje a Héctor Pucciarelli, pp. 87-109. Puerto Madryn, CONICET.
Neves, W.A. & Pucciarelli, H.M. (1991) “Morphological affinities of the first Americans: an exploratory analysis based on early south American human remains.” Journal of Human Evolution 21(4), pp. 261-273.
Neves, W.A. & Hubbe, M. (2005) “Cranial morphology of early Americans from Lagoa Santa, Brazil: Implications for the settlement of the New World.” Proceedings of the National Academy of Sciences 102(51), pp. 18309-18314.
Newman, M.T. (1943) “A metric study of undeformed Indian crania from Peru.” American Journal of Physical Anthropology 1(1): 21-45.
Olson, D.M., Dinerstein, E., Wikramanayake, E.D., Burgess, N.D., Powell, G.V.N., Underwood, E.C., Damico, J.A., Ituoa, I., Strand, H.E., Morrison, J.C., Loucks, C.J., Allnutt, T.F., Ricketts, T.H., Kura, Y., Lamoreux, J.F., Wettengel, W.W., Hedao, P. & Kassem, K. R.(2001) “Terrestrial ecoregions of the world: a new map of life on earth”. BioScience 5(11), pp. 933-938.
Paschetta, C., de Azevedo, S., Castillo, L., Martínez‐Abadías, N., Hernández, M., Lieberman, D. E. & González‐José, R. (2010) “The influence of masticatory loading on craniofacial morphology: A test case across technological transitions in the Ohio valley.” American Journal of Physical Anthropology 141(2), pp. 297-314.
Paschetta, C., González-José, R. & Lanata, J.L. (2017) De Cómo Cruzar Fronteras en la Ciencia. Homenaje a Héctor M. Pucciarelli. Puerto Madryn y San Carlos de Bariloche, IPCSH e IIDyPCa, CONICET.
Pérez, S.I. & Monteiro, L.R. (2009) “Non-random factors in modern human morphological diversification: A study of craniofacial variation in southern south American populations.” Evolution 63(4), pp. 978-993.
Pérez, S.I., Bernal, V., Gonzalez, P.N., Sardi, M. & Politis, G.G. (2009) “Discrepancy between cranial and DNA data of early Americans: Implications for American peopling.” PLoS ONE 4(5), pp. e5746.
Pérez, S.I., Lema, V., Diniz‐Filho, J.A. F., Bernal, V., Gonzalez, P.N., Gobbo, D., & Pucciarelli, H.M. (2011) “The role of diet and temperature in shaping cranial diversification of South American human populations: an approach based on spatial regression and divergence rate tests.” Journal of Biogeography 38(1), pp. 148-163.
Pillai, K.C.S. (1977) “Distribution of characteristic roots in multivariate analysis. Part II. Non‐Null Distribution.” Canadian Journal of Statistics 5, pp. 1-62.
Ponce de León, M.S., Koesbardiati, T., Weissmann, J.D., Milella, M., Reyna-Blanco, C.S., Suwa, G., Kondo, O., Malaspinas, A.S., White, T.D. & Zollikofer, C.P.E. (2018) “Human bony labyrinth is an indicator of population history and dispersal from Africa.” Proceedings of the National Academy of Sciences 115(16), pp. 4128-4133.
Posth, C., Nakatsuka, N., Lazaridis, I., Skoglund, P., Mallick, S., Lamnidis, T.C., Rohland, N., Nagele, K., Adamski, N., Bertolini, E., Broomandkhoshbacht, N., Cooper, A., Culleton, B.J., Ferraz, T., Ferry, M., Furtwängler, A., Haak, W., Harkins, K., Harper, T.K., Hünemeier, T., Lawson, A.M., Llamas, B., Michel, M., Nelson, E., Oppenheimer, J., Patterson, N., Schiffels, S., Sedig, J., Stewardson, K., Talamo, S., Wang, C.C., Hublin, J.J., Hubbe, M., Harvati, K., Nuevo Delaunay, A., Beier, J., Francken, M., Kaulicke, P., Reyes-Centeno, H., Rademaker, K., Trask, W.R., Robinson, M., Gutierrez, S.M., Prufer, K.M., Salazar-García, D.C., Chim, E.N., Müller Plumm Gomes, L., Alves, M.L., Liryo, A., Inglez, M., Oliveira, R.E., Bernardo, D.V., Barioni, A., Wesolowski, V., Scheifler, N.A., Rivera, M.A., Plens, C.R., Messineo, P.G., Figuti, L., Corach, D., Scabuzzo, C., Eggers, S., DeBlasis, P., Reindel, M., Méndez, C., Politis, G., Tomasto-Cagigao, E., Kennett, D.J., Strauss, A., Fehren-Schmitz, L., Krause, J. & Reich, D. (2018) “Reconstructing the Deep Population History of Central and South America”. Cell 175(5), pp.1185-1197.
Pucciarelli, H.M. (2008) Evolución y Diversificación Biológica Humana desde la Perspectiva Craneofuncional. México D.F., Universidad Autónoma de México.
Pucciarelli, H.M., Sardi, M.L., López, J.C.J. & Sanchez, C.S. (2003) “Early peopling and evolutionary diversification in America.” Quaternary International 109, pp. 123-132.
Pucciarelli, H.M., Neves, W.A., González-José, R., Sardi, M.L., Ramírez Rozzi, F., Struck, A. & Bonilla, M.Y. (2006) “East-west cranial differentitation in pre-Columbian populations of South America.” Homo 54(3), pp. 133-150.
Pucciarelli, H.M., Gonzalez-José, R., Neves, W.A., Sardi, M. & Ramírez Rozzi, F. (2008) “East-west cranial differentiation in pre-Columbian populations from Central and North America.” Journal of Human Evolution 54(3), pp. 296-308.
Pucciarelli, H.M., Pérez, S.I. & Politis, G.G. (2010) “Early Holocene human remains from the Argentinean pampas: additional evidence for distinctive cranial morphology of early south Americans.” American Journal of Physical Anthropology 143(2), pp. 298–305.
R Core Team (2013) R: A language and environment for statistical computing. R Foundation for Statistical Computing. Vienna.
Rademaker, K. & Hodgins, G. (2018) “Exploring the chronology of occupations and burials at Cuncaicha rockshelter, Peru.” En: Harvati, K., Jaeger, G. & Reyes-Centeno, H. (eds.) New Perspectives on the Peopling of the Americas, Kerns Verlag, Tuebingen, pp. 107-124.
Rademaker, K., Hodgins, G., Moore, K., Zarrillo, S., Miller, C., Bromley, G.R.M., Leach, P., Reid, D.A., Yépez Álvarez, W. & Sandweiss, D.H. (2014) “Paleoindian settlement of the high-altitude Peruvian Andes.” Science 346, pp. 466-469.
Ross, A.H., Ubelaker, D.H. & Guillén, S. (2008) “Craniometric patterning within ancient Peru.” Latin American Antiquity 19(2), pp. 158-166.
Ross, A.H. & Ubelaker, D.H. (2019) “Complex Nature of Hominin Dispersals: Ecogeographical and Climatic Evidence for Pre-Contact Craniofacial Variation.” Scientific Reports 9, pp. 11743.
Sardi, M.L. (2017) “El método craneofuncional.” En: Paschetta, C., González-José, R. & Lanata, J.L. (eds.) De cómo cruzar fronteras en la ciencia. Homenaje a Héctor Pucciarelli, Puerto Madryn CONICET, pp. 120-135.
Sardi, M.L., Ramírez-Rozzi, F., Dahinten, S. & Pucciarelli, H.M. (2004) “Amerindians: testing the hypothesis about their homogeneity.” Comptes Rendus Palevol 3(5), pp. 403-409.
Sardi, M.L., Ramírez-Rozzi, F., González-José, R. & Pucciarelli, H.M. (2005) “South Amerindian craniofacial morphology: diversity and implications for Amerindian evolution”. American Journal of Physical Anthropology 128(4), pp. 747–766.
Sardi, M.L., Novellino, P.S. & Pucciarelli, H.M. (2006) “Craniofacial morphology in the Argentine Center‐West: consequences of the transition to food production.” American Journal of Physical Anthropology 130(3), pp. 333-343.
Shrout, P.E. & Fleiss, J.L. (1979) “Intraclass correlations: uses in assessing rater reliability.” Psychological Bulletin 86(2), p. 420.
Stone, J.H., Chew, K., Ross, A.H. & Verano, J.W. (2015) “Craniofacial plasticity in ancient Peru.” Anthropologischer Anzeiger 72(2), pp. 169-183.
Tarazona-Santos, E., Carvalho-Silva, D.R., Pettener, D., Luiselli, D., De Stefano, G.F., Martinez Labarga, C., Rickards, O., Tyler-Smith, C., Pena, S.D.J. & Santos, F.R. (2001) “Genetic differentiation in Amerindians is related to environmental and cultural diversity: evidence from the Y chromosome.” American Journal of Human Genetics 68(6), pp. 1485-1496.
Verano, J. (1987) “Cranial microvariation at Pacatnamu: a study of cementery population variability.” Tesis Doctoral. University of California.
von Cramon-Taubadel, N., Strauss, A. & Hubbe, M. (2017) “Evolutionary population history of early Paleoamerican cranial morphology.” Science Advances 3(2), p. e1602289.
Walker, M.J., Berkelhammer, M., Björck, S., Cwynar, L.C., Fisher, D.A., Long, A.J., Lowe, J.J, Newnham, R., Rasmussen, S.O. & Weiss, H. (2012) “Formal subdivision of the Holocene Series/Epoch: a discussion paper by a working group of INTIMATE (Integration of ice core, marine and terrestrial records) and the subcommission on Quaternary Stratigraphy (International Commission on Stratigraphy).” Journal of Quaternary Sciences 27(7), pp. 649-659.
Ward, J.H. (1963) “Hierarchical grouping to optimize an objective function.” Journal of the American Statistical Association 58(301), pp. 236-244.
Zar, J.H. (2010) Biostatistical Analysis. Nueva Jersey, Prentice Hall.
Menéndez, L.P., Sardi, M.L., Scheifler, N.A., González, M.E., Messineo, P. & Politis, G.G. (2019a) “10,000 years of mandibular evolution in southern South America: Implications for morphological diversification.” American Journal of Physical Anthropology 168, pp. 164
Menéndez, L.P., Rademaker, K & Harvati, K. (2019b) “Revisiting east-west skull patterns and the role of random factors in south America: cranial reconstruction and morphometric analysis of the facial skeleton from Cuncaicha rock shelter (southern Peru).” PaleoAmerica 5(4), pp. 315-334

