Resumen: Se analiza aquí, la influencia del rociado y vigilancia sobre la estructuración de las poblaciones peri e intra-domésticas de T. infestans usando morfología cuantitativa. Las colectas en intra (ID) y peridomicilio (PD) se realizaron sobre una población libre de rociado por 10 años y en poblaciones hasta 8 años post rociado (población con vigilancia y rociado). En prerociado, no se observó estructuración poblacional entre ID y PD y 6 de 7 domicilios mostraban infestación (40 adultos de T. infestans). Post intervención, se detectaron 6 adultos de T. infestans en ID. Estos insectos mostraron similitud morfológica con T. infestans residuales. Los resultados plantean que en presencia de vigilancia entomológica, los ID pueden permanecer libres de T. infestans aún con PD infestados.
Palabras clave: Triatoma infestans, morfometría, sensilla antenales, enfermedad de Chagas.
Abstract: The influence of spraying and surveillance on the structuring of peri and intra-domestic populations of T. infestans is analyzed here using quantitative morphology. The collections in intra (ID) and peridomicilio (PD) were carried out on a population free of spraying for 10 years and in populations up to 8 years post sprayed (population with surveillance and spraying). In pre-spraying, no population structuring was detected between ID and PD. Post intervention, 6 adults of T. infestans were detected in ID. These insects showed morphological similarity with residual T. infestans. The results suggest that in the presence of entomological surveillance, IDs can remain free of T. infestans even with infested PD.
Keywords: Triatoma infestans, morphometry, antennal sensilla, Chagas disease.
EFECTO DE LA VIGILANCIA ENTOMOLÓGICA EN LA ESTRUCTURACIÓN POBLACIONAL DE Triatoma infestans (HEMIPTERA, REDUVIIDAE) DE INTRA Y PERIDOMICILIO EN UN ÁREA ENDÉMICA DE LA PROVINCIA DE LA RIOJA (ARGENTINA)
Recepción: 15 Abril 2020
Aprobación: 13 Julio 2020
En Argentina, Triatoma infestans es el principal vector del parásito Trypanosoma cruzi, agente causal de la enfermedad de Chagas. En la provincia de la Rioja, éste insecto se encuentra presente en viviendas rurales y desarrolla importantes colonias en ambientes peridomésticos muy cercanos a los domicilios. Las poblaciones peridomésticas de T. infestans plantean serios problemas para su eliminación. Ciertos hábitats como gallineros o corrales de cabras, resultan excelentes refugios para triatominos y dificultan la llegada del insecticida. Además, luego del rociado, los insecticidas quedan expuestos a la degradación ambiental, limitando su eficacia y aumentando la probabilidad de permanencia de poblaciones residuales. Los insectos peridomésticos residuales a un rociado pueden servir como fuente de reinfestación al dispersarse hacia los domicilios cercanos. En triatominos, la dispersión activa es clave tanto para la colonización de nuevos hábitats como para la reinfestación, luego de un rociado con insecticidas. Schofield et al. (1999) plantearon que el vuelo es una actividad importante para la búsqueda y colonización de hábitats en T. infestans. La información existente en cuanto a la actividad de vuelo en T. infestans según el sexo muestra discrepancias. Ciertos autores plantean a las hembras con mayor actividad dispersiva por vuelo (Gurevitz et al., 2006; Lehane & Schofield, 1982) mientras que en otros casos son los machos quienes muestran mayor actividad de vuelo (Ceballos et al., 2005; Vazquez-Prokopec et al., 2006). Aún, Lobbia et al. (2019) proponen que la resistencia a insecticidas reduce la capacidad dispersiva en hembras de T. infestans. Abrahan et al. (2011) demostraron que la dispersión activa caminando es también una estrategia empleada por T. infestans especialmente por las hembras que transportan numerosa cantidad de huevos, o por adultos con buen estado nutricional, que no podrían dispersarse por vuelo.
En Argentina, a pesar de los esfuerzos y las acciones de control vectorial, T. infestans todavía representa un importante problema de salud pública (OPS, 2006). En áreas históricamente endémicas, cuando no se desarrollan acciones de control vectorial por períodos mayores a 10 años, se ha observado una alta infestación en el interior de las viviendas (intradomicilio). Existen casos donde el intradomicilio presenta baja o nula infestación por T. infestans con peridomicilios infestados a pocos metros de las viviendas. Estos datos, plantean hipótesis contradictorias sobre la importancia de los peridomicilios infestados en la probabilidad de reinfestación del intradomicilio y la frecuencia necesaria de evaluación y rociado que garantice que los intradomicilios se mantengan libres del vector. Cada hábitat presenta características específicas en cuanto al número y tipo de hospedador, condiciones ambientales locales, permanencia de los hospedadores, etc. Todas estas particularidades del hábitat, parecen afectar de un modo específico las características morfofisiológicas de T. infestans resultando en cambios que serían de utilidad para la identificación de insectos recolonizantes o con diferentes capacidades dispersivas (Schachter-Broide et al., 2004; Abrahan et al., 2008; Hernández et al., 2011; Hernández et al., 2013). Diversas son las investigaciones que dan cuenta del uso la técnica de morfometría geométrica y fenotipos antenales en triatominos para análisis de dispersantes (Dumonteil et al., 2007), estructura poblacional (Vargas et al., 2006; Schachter-Broide et al., 2004; Hernández et al., 2011; Gaspe et al., 2012) características fenéticas compatibles con vuelo, entre otros (Hernández et al., 2015; Hernández et al., 2020). El análisis de la estructuración espacial poblacional permite determinar la continuidad que puede existir en la distribución geográfica de las poblaciones de una especie, mediante la caracterización de sus individuos y la comparación de variables relacionadas al fenotipo, al genotipo, o ambas (Dujardin et al., 2002).
Este tipo de estudio es importante a la hora de definir las estrategias de control vectorial ya que si las poblaciones están aisladas o altamente estructuradas, tendrían menor intercambio de individuos con poblaciones vecinas, lo que haría menos probable que los hábitats ya tratados vuelvan a ser re-invadidos y/o re-colonizados y que nuevos hábitats se tornen positivos. Las variables morfológicas resultan de utilidad para el estudio de la reinfestación y estructuración poblacional, toda vez que poblaciones morfológicamente diferentes estarían espacialmente estructuradas, y presentarían bajo o nulo movimiento dispersivo entre ellas. Se analiza aquí la influencia de la frecuencia de evaluación entomológica y rociado con insecticidas sobre la estructuración de las poblaciones peri e intradomésticas de T. infestans en dos localidades del Dpto. Independencia (La Rioja), usando morfología cuantitativa (morfometría geométrica de cabezas y fenotipos antenales). Se plantea la hipótesis de que las poblaciones sin rociado y sin vigilancia mostrarán similitud fenotípica entre intra y peridomicilio debido a que existe un mayor flujo entre estos dos hábitats. Contrariamente, luego del rociado y debido a la vigilancia entomológica, se espera que el número de T. infestans que lleguen a un domicilio y logren colonizarlo sea bajo o nulo resultando en poblaciones de intra y peridomicilio fenotípicamente diferentes.
El departamento Independencia es parte de Los Llanos riojanos, la región de La Rioja con mayor pobreza, aridez y mayores infestaciones dentro de la provincia (Porcasi et al., 2007; Cavallo et al., 2018). Las viviendas rurales características son construcciones de adobe y techos de material vegetal principalmente. Rodeando el área domiciliar se encuentra el peridomicilio que puede abarcar desde unos pocos metros hasta los 80 metros alrededor del domicilio. La cría de animales domésticos es muy común en esta región, siendo los corrales de cabras y gallineros los principales hábitats que rodean los domicilios. Al iniciar el estudio (2006) el sitio de estudio llevaba 10 años sin ningún tipo de tratamiento con insecticidas (Porcasi et al. 2007), por lo que las poblaciones de T. infestans colectadas en esta fecha pueden considerarse como si nunca hubieran sido tratadas. A finales de 2006 y en 2009, el Programa Provincial de Chagas (PPCh) realizó rociado masivo con insecticidas piretroides en intra y peridomicilio.
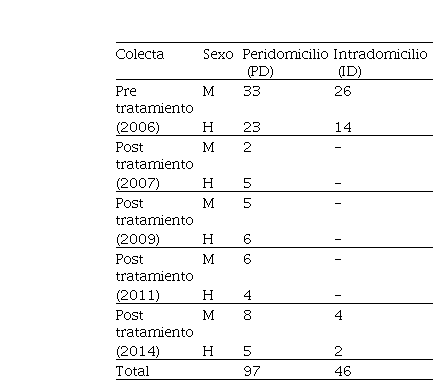
Tabla 1: Número de ejemplares colectados en intra y peridomicilio en pre tratamiento (2006) y post tratamiento con insecticidas
En el interior de las viviendas se realizó el rociado tradicional con betacipermetrina. En el peridomicilio, el tratamiento no fue el habitual, sino que consistió en la aplicación de cipermetrina (6%) en cabras y gallinas (5 y 1 cm3 respectivamente) y fipronil en perros (5 cm3, al 1%) colocado como pouron en la base del cuello. Luego de los rociados, el Programa de Chagas realizó vigilancia entomológica (con una frecuencia variable según la disponibilidad de presupuesto del PPCh) y efectuó tratamiento local cuando se reportó un intradomicilio positivo. Las colectas de T. infestans en intra y peridomicilio fueron realizadas por los investigadores por el método de búsqueda activa (hora/hombre) usando tetrametrina 0,2% como desalojante (Spacial 0,2, provisto por el Ministerio de Salud de La Nación). Se inspeccionaron 7 viviendas, 8 corrales de cabras y 8 gallineros en octubre de 2006 (población sin rociado ni vigilancia entomológica previa) y en enero, abril y octubre de 2007, septiembre de 2009, octubre de 2011 y marzo de 2014 (poblaciones con vigilancia y rociado).
Las viviendas seleccionadas para este estudio correspondieron a una de las zonas de mayor infestación en intra y peridomicilio en el momento de la revisión previa al tratamiento con insecticidas y mayores denuncias de pobladores por la presencia de triatominos en el interior de sus viviendas en post rociado. Los adultos de T. infestans colectados en estas fechas fueron separados para los estudios morfológicos. Se analizaron 143 adultos de T. infestans de las localidades de Salinas de Bustos (S 30,2638 W 67,606367, 1128 msnm) y La Torre (S 30,20346 W 67,60602, 1182 msnm),(Tabla 1), distantes 7 Km una de la otra. Los insectos fueron llevados al Laboratorio de Entomología (CRILAR-CONICET, La Rioja, Argentina) donde se separaron las cabezas y las antenas para su posterior análisis morfológico. Las antenas y cabezas fueron rotuladas y conservadas en alcohol 70% para su posterior análisis. Para el conteo de sensilla antenales,se utilizó la técnica descripta por Catalá y Schofield (Catalá & Schofield, 1994). Se identificaron y contaron 3 quimiosensilla de tipo tricoide (tricoide de pared fina del pedicelo y flagelo 1 y tricoide de pared gruesa del flagelo 1) y un mecanoreceptor (Bristles del pedicelo). Para morfometría geométrica, se fotografiaron las cabezas montadas sobre un soporte metálico usando un esteromicroscopio (10X).
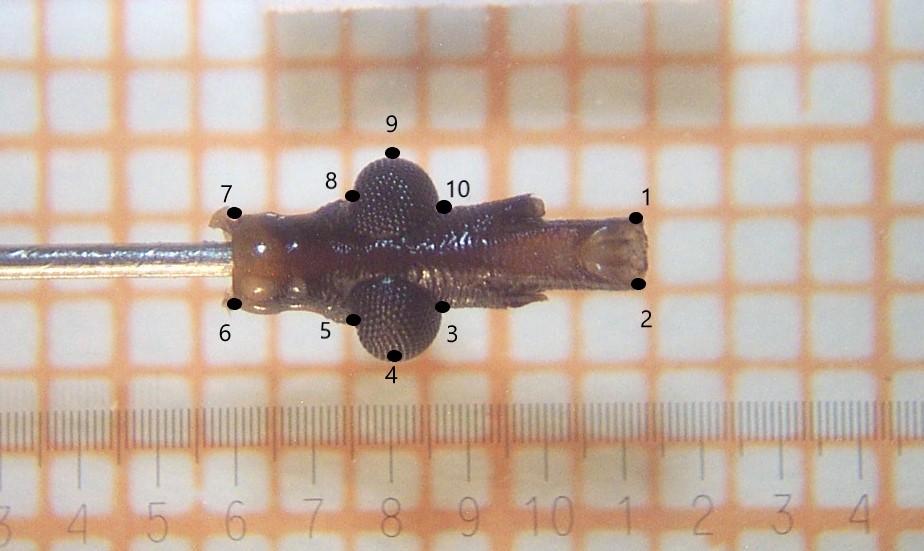
Figura 1
Figura 1: Vista ventral de la cabeza de Triatoma infestans con los 10 landmarks tipo II tomados para los análisis morfométricos. Los números indican el orden de captura de los landmarks. Fuente: Elaboración propi
Sobre las fotos se marcaron 10 landmarks tipo II (Figura 1). La obtención de las variables de tamaño (centroid size) y conformación (partial warps) se realizó por medio del Análisis generalizado de Procrustes (AGP). La colecta de landmarks, la obtención de variables de tamaño y conformación, así como los análisis multivariados con las sensilla antenales se realizaron utilizando el módulo CLIC 49 (http://xyom-clic.eu/the-clic-package/). Las variables de conformación derivadas del AGP, fueron utilizadas para el análisis discriminante. La significancia en el análisis discriminante fue evaluada usando los valores de Wilks y las distancias de Mahalanobis. Estos valores fueron evaluados usando test no paramétricos basados en permutaciones (5000 runs). La corrección de Bonferroni fue usada para comparaciones múltiples. El análisis de similitud fenotípica fue realizado entre poblaciones de T. infestans de pre y post rociado con insecticidas. Para dicho análisis se utilizó el test cross-check del software CLIC 49. Éste es un test no paramétrico que valida la reclasificación del análisis discriminante a través de permutaciones (5000 runs) asignando a cada individuo el grupo con el que comparte mayor similitude fenotípica.
En pre tratamiento con insecticidas, 6 de los 7 domicilios (85,7%) y todos los hábitats peridomésticos resultaron positivos para T. infestans. En post tratamiento con insecticidas se registraron dos domicilios positivos en 2014 (28,6%), mientras que en 2007, 2009 y 2011 los intradomicilios se mantuvieron libres del vector. El número de insectos colectados por hábitat y fecha se resume en la Tabla 1.
En 2006, cuando el departamento llevaba 10 años sin tratamiento, no se observó estructuración poblacional entre intra y peridomicilio usando variables antenales y de cabezas.
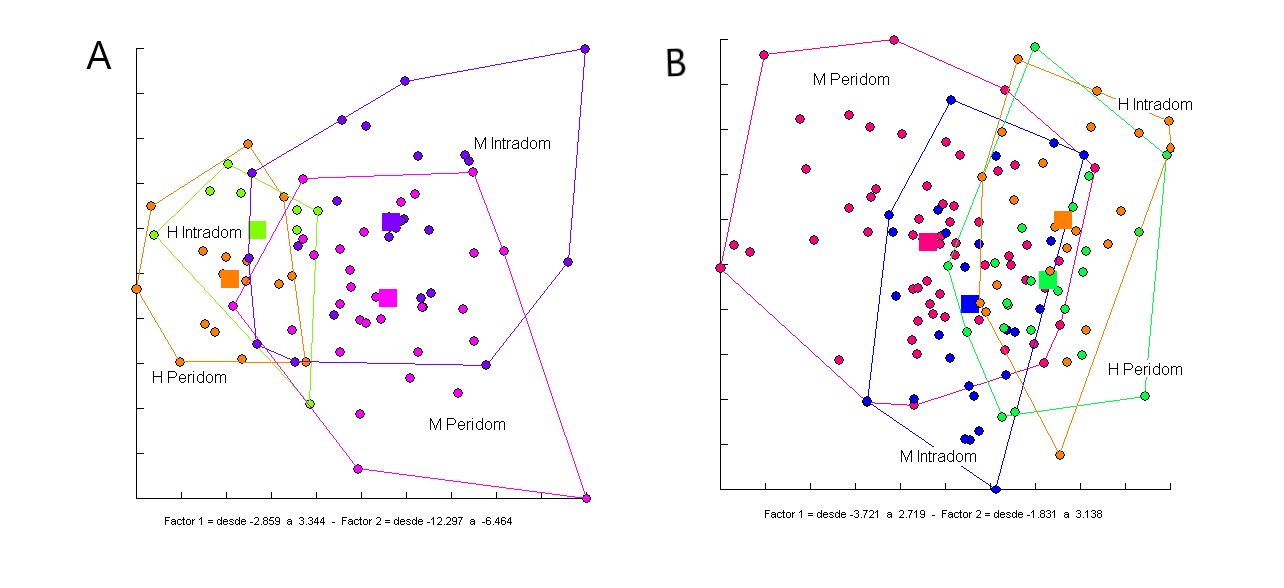
Figura 2
Figura 2: Análisis discriminante con las 4 variables antenales (A) y con las variables de conformación de la cabeza (B) de T. infestans colectados en pre tratamiento con insecticidas (2006) en intra y perdidomicilio. Peridom: Peridomicilio, Intradom: Intradomicilio, H: Hembras, M: Machos. Fuente: Elaboración propia.
El análisis multivariado discriminante con 4 variables de la antena no encontró diferencias entre intra y peridomicilio, aunque si diferencias sexuales (Valor p < 0,001). El tamaño centroide de cabezas no difirió entre ejemplares de intra y peridomicilio en ninguno de los sexos (CS machos ID: 354,34, machos PD: 355,11, hembras ID: 368,23, hembras PD: 369,21). La conformación de la cabeza no mostró diferencias entre hábitats eliminando el efecto alométrico (p > 0,001) (Figura 2). Entre 2007 y 2011 las colectas en intradomici-lios fueron negativas para T. infestans en las dos localidades muestreadas. En la colecta de 2014 se detectó la presencia de 6 adultos (4 machos y 2 hembras) de T. infestans dentro de uno de los intradomicilios muestreados. Debido al pequeño tamaño de la muestra colectada en intradomicilio en post tratamiento con insecticida, no se pudo realizar un análisis discriminante para el análisis de la estructuración poblacional. Para esto, se realizó un análisis de similitud fenotípica, en el cual por medio de un test de permutaciones (5000 ciclos), los individuos de intradomicilio son asignados a los grupos de intra y al peridomicilio para evaluar con cual comparte mayor similitud fenotípica en base a fenotipos antenales y a conformación de la cabeza. Los análisis de similitud fenotípica mostraron que 4/6 (66,67%) mostraron fenotipos antenales y conformación de cabeza compatibles con T. infestans de intradomicilio y el 33,33%restante (2/6) mostraron fenotipos compatibles con insectos de peridomicilio.
Elaboración propia.Los resultados sugieren que el tratamiento con insecticidas y la vigilancia entomológica tienen un efecto sobre las poblaciones de ID y PD de Triatoma infestans. Los resultados expuestos plantean que en presencia de vigilancia entomológica los ID pueden permanecer libres de T. infestans aún con PD infestados. Luego de 10 años de cese de las acciones sistemáticas de control contra T. infestans en Los Llanos riojanos y particularmente en el departamento Independencia, las poblaciones de este vector pueden considerarse como si nunca hubieran recibido tratamiento con insecticidas piretroides. Diversas investigaciones indican que las poblaciones se recuperan con cierta facilidad (en 3 a 4 años) de un tratamiento insecticida, a partir de focos residuales, sobre todo en peridomicilios complejos y con buen acceso a animales de cría como fuente alimenticia (Cecere et al., 2004), desde donde también pueden dispersarse hacia hábitats vecinos (Abrahan et al., 2011; Abrahan et al., 2016; Vazquez-Prokopec et al., 2004). El análisis de las características de las poblaciones en esta fase previa al rociado (línea base) es importante para comprender cómo se comporta T. infestans en una situación en la que no ha existido control tradicional con insecticidas, y observar posteriormente la influencia de la aplicación del insecticida y la vigilancia entomológica en dicha población. Las características fenotípicas morfológicas y de una población son informativas acerca de la estructura poblacional y de la movilidad relativa de su individuos hacia ambientes cercanos (Schachter-Broide et al. 2004, Abrahan et al. 2008, Hernández et al 2011). La similitud fenotípica entre T. infestans de ID y PD en el prerociado podría indicar un elevado intercambio de insectos entre éstos hábitats posiblemente en busca de fuentes de alimento debido a un incremento de las poblaciones. López et al. (1999) encontraron que un 30% de los T. infestans de un gallinero cercano a una vivienda presentaban sangre humana en sus intestinos, lo cual hace pensar sobre el movimiento de estos insectos entre hábitats próximos en busca de alimento. La dispersión caminando entre hábitats cercanos podría ser una estrategia de llegar a un nuevo hospedador donde alimentarse, quizás sin el objetivo de colonizar el hábitat, sino como parte de su búsqueda habitual de una fuente de sangre, dentro de cortas distancias. En el momento de comenzar el estudio, el departamento contaba con una infestación de 45,1% en intradomicilio y 57,3% en peridomicilio (Porcasi et al., 2007). Luego de 3 años, la infestación fue de 16,25% y 27,5% para intra y peridomicilio respectivamente (Programa Provincial de Chagas, 2011, no publicado), debido a las acciones de control realizadas en el departamento. Los resultados de este trabajo muestran que el rociado tradicional eliminó las poblaciones intradomésticas de T. infestans de las viviendas muestreadas, logrando mantener las habitaciones humanas libres de este insecto durante los siguientes 3 años, luego del cual la infestación intradomiciliaria detectada fue muy baja (solo uno de los ID fue positivo). Teniendo en cuenta que los propios pobladores y el Programa Provincial de Chagas actuaban como agentes de vigilancia (los domicilios eran tratados cada vez que se denunciaba la presencia de triatominos en el interior de una vivienda), se asume que estas acciones mantuvieron los intradomicilios libres de colonizaciones por T. infestans en el período de tiempo analizado. Estos datos confirman el bajo movimiento de T. infestans entre intra y peridomicilio en una zona de alto endemismo del vector. Es probable que esta especie prefiera dispersarse entre hábitats peridomésticos más que hacia el interior de la vivienda humana en busca de sangre. Las habitaciones humanas podrían ser hábitats menos atractivos para T. infestans. Catalá (2011) plantea que los corrales de cabras serían los hábitats más atrayentes para los dispersantes de esta especie debido a la mayor radiación infrarroja emitida desde este hábitat, dada por el calor corporal de los numerosos hospedadores. El bajo intercambio de T. infestans entre peri e intradomicilio tiene importantes implicancias en la transmisión de la enfermedad de Chagas. En otros dos departamentos riojanos (San Martín y Castro Barros) la transmisión vectorial de la enfermedad pudo interrumpirse aún con infestaciones peridomésticas de 25 − 30% (Gorla et al., 2010). Esto permite postular que no sería necesario buscar la eliminación total de las poblaciones peridomésticas de T. infestans a corto plazo (teniendo en cuenta los presupuestos acotados con los que cuentan muchas veces los programas de control, vectorial), sino apuntar a un control y vigilancia de las mismas y a un mejoramiento del intradomicilio que impida nuevas colonizaciones de las habitaciones humanas. La idea de un bajo movimiento de T. infestans hacia el intradomicilio, es un dato relevante para el control vectorial ya que indica que si se refuerza la vigilancia en los intradomicilios con participación comunitaria, educación, información y mejoramiento de la vivienda la transmisión vectorial debería tender a cero aunque persistan focos peridomiciliarios de T. infestans. Futuras investigaciones en éste y otros departamentos con alta infestación de la provincia, podrían permitirnos ampliar las conclusiones abordadas en cuanto al efecto de la vigilancia entomológica en la estructuración poblacional de T. infestans. Además permitirían discutir sobre posibles efectos de la zona y/o de la utilización del insecticida, que ayudarían a sugerir recomendaciones para ser aplicadas en las campañas de control de triatominos de La Rioja.
Agradecimientos
A Ivana Amelotti, Luciana Abrahan y Carlos Nieto por la ayuda en las colectas de campo. Proyecto financiado por el PICT 2883. MLH es miembro de la Carrera de Investigador Científico de CONICET (Argentina).
Tabla 1: Número de ejemplares colectados en intra y peridomicilio en pre tratamiento (2006) y post tratamiento con insecticidas
Figura 1
Figura 1: Vista ventral de la cabeza de Triatoma infestans con los 10 landmarks tipo II tomados para los análisis morfométricos. Los números indican el orden de captura de los landmarks. Fuente: Elaboración propi
Sobre las fotos se marcaron 10 landmarks tipo II (Figura 1). La obtención de las variables de tamaño (centroid size) y conformación (partial warps) se realizó por medio del Análisis generalizado de Procrustes (AGP). La colecta de landmarks, la obtención de variables de tamaño y conformación, así como los análisis multivariados con las sensilla antenales se realizaron utilizando el módulo CLIC 49 (http://xyom-clic.eu/the-clic-package/). Las variables de conformación derivadas del AGP, fueron utilizadas para el análisis discriminante. La significancia en el análisis discriminante fue evaluada usando los valores de Wilks y las distancias de Mahalanobis. Estos valores fueron evaluados usando test no paramétricos basados en permutaciones (5000 runs). La corrección de Bonferroni fue usada para comparaciones múltiples. El análisis de similitud fenotípica fue realizado entre poblaciones de T. infestans de pre y post rociado con insecticidas. Para dicho análisis se utilizó el test cross-check del software CLIC 49. Éste es un test no paramétrico que valida la reclasificación del análisis discriminante a través de permutaciones (5000 runs) asignando a cada individuo el grupo con el que comparte mayor similitude fenotípica.
Figura 2
Figura 2: Análisis discriminante con las 4 variables antenales (A) y con las variables de conformación de la cabeza (B) de T. infestans colectados en pre tratamiento con insecticidas (2006) en intra y perdidomicilio. Peridom: Peridomicilio, Intradom: Intradomicilio, H: Hembras, M: Machos. Fuente: Elaboración propia.
El análisis multivariado discriminante con 4 variables de la antena no encontró diferencias entre intra y peridomicilio, aunque si diferencias sexuales (Valor p < 0,001). El tamaño centroide de cabezas no difirió entre ejemplares de intra y peridomicilio en ninguno de los sexos (CS machos ID: 354,34, machos PD: 355,11, hembras ID: 368,23, hembras PD: 369,21). La conformación de la cabeza no mostró diferencias entre hábitats eliminando el efecto alométrico (p > 0,001) (Figura 2). Entre 2007 y 2011 las colectas en intradomici-lios fueron negativas para T. infestans en las dos localidades muestreadas. En la colecta de 2014 se detectó la presencia de 6 adultos (4 machos y 2 hembras) de T. infestans dentro de uno de los intradomicilios muestreados. Debido al pequeño tamaño de la muestra colectada en intradomicilio en post tratamiento con insecticida, no se pudo realizar un análisis discriminante para el análisis de la estructuración poblacional. Para esto, se realizó un análisis de similitud fenotípica, en el cual por medio de un test de permutaciones (5000 ciclos), los individuos de intradomicilio son asignados a los grupos de intra y al peridomicilio para evaluar con cual comparte mayor similitud fenotípica en base a fenotipos antenales y a conformación de la cabeza. Los análisis de similitud fenotípica mostraron que 4/6 (66,67%) mostraron fenotipos antenales y conformación de cabeza compatibles con T. infestans de intradomicilio y el 33,33%restante (2/6) mostraron fenotipos compatibles con insectos de peridomicilio.
Elaboración propia.

